BIO254:PlaceCells

What Are Place Cells
Place cells are neurons found in the hippocampus that trigger an action potential when an animal is in a certain location within an environment. Thus, these cells constitute a "place-field" and are thought to be pyramidal cells (excitatory neurons) in the CA1 and CA3 regions of hippocampus. Several studies confirmed that place cells reside in the hippocampus, because when the hippocampus was removed or destroyed in rats, they display significantly impaired place navigation when swimming through the Morris water maze. Granule cells in the dentate gyrus of hippocampus are also believed to be place cells.
O'Keefe and Dostrovsky were the first scientists to describe the phenomenon of place cells. From their observations, they hypothesized that the (rat) hippocampus serves to form a cognitive map of the environment. Interestingly, Ekstrom et al found cells with similar properties of place cells in the human hippocampus by doing extracellular recordings from epileptic children.
Place Cells As Pointers in the Cognitive Map
When investigations of animal learning first began, advocates of the dominant "stimulus-response" (SR) theory argued that maze learning is achieved by direct associations between stimuli and rewarded behavioral responses. In 1948, however, Tolman demonstrated that rats can actually navigate mazes using shortcuts, suggesting that attributes of rat learning behavior could not necessarily be explained by SR theory. As a result of his experiments, Tolman concluded that rats may create and then utilize cognitive maps--or global representations of the environment--to localize directional cues in mazes.
The proposal of cognitive maps was first critiques for its lack of compelling evidence for an underlying neural mechanism. Upon the discovery of place cells by O'Keefe and Dostrovsky, scientists began to think that these neurons might be the building blocks of the long-hypothesized cognitive map.
As a rat explores the environment at large, we can readily correlate significant increases in a place cell's firing rate with the specific location of the animal. The firing rate can exceed 100 Hz or not even fire at all. O'Keefe and colleagues observed that hippocampal neurons even maintained their spatial firing pattern when certain environmental cues used in maze tests were removed, but not when the shape of the environment was altered. This indicated that there is a threshold of cues required for rats to sufficiently define their "cognitive maps."
What's in a Place Cell "Map"?
One problem with the place map hypothesis is the lack of evidence indicating how place cells are able to communicate one's "location" with respect to "other locations." Likewise, how many place cells are required to create a cognitive map? To identify one's location in an environment requies that the entire area be represented, which would be most simply represented by place fields that are uniformly distributed throughout the environment. In contrast to this logic, early studies show that "space" in the hippocampus is not represented in this fashion; place fields are not continuous or topographically organized. Rather, there is evidence that place fields are made up of clusters of neighboring cells. It seems that hippocampal representation does not involve a homogeneous representation of the entire area of physical space.
Place Cells, sometimes called Space Cells are a collection of neurons, found in the Hippocampus, that activate in response to an animal's position in it's physical environment. Together, a set of place cells form an abstract map of the geometry of an animal's environment. Each Place Cell has a place field, or mapping of areas to which it is most sensitive to. The firing of many different place cells, according to their place fields and the physical postion of the animal, specify the animals location and possibly it's orientation. When an animal is placed in a new environment, it's cognitive spatial map is likely to be quite disorganized. Experimentaly, this can be observed as the erratic firing of place cells, without regard to the physical location of the animal. However, the place fields could be established in minutes after the initial exposure to a new environment. A typical experiment might consist of a freely moving rodent implanted with extracellular electrodes and placed in a novel enclosure. In this way, the firing of many place cells can be recorded in conjunction with the animals location. Over time, an organized map with distinct place fields for different cells will emerge. Indeed, it has been found that through repeated exposures animals can progressively separate and fine tune their representations of two similar environments. Once they are tuned by experience, these spatial representations are robust and can remain stable for months.
When an animal is placed in a different environment, a place cell could have completely different place field, or even no place field. This is the so called remapping. Indeed, in any given environment, only 40% of place cells will be active.
The precise mechanisms of plasticity that tune and maintain the cognitive spatial map are not known. However, it is likely that well studied mechanisms of plasticity such as LTP and LTD are involved. Animals in which molecules involved in LTP and LTD (NMDAR, CaMKII, etc.) have been perturbed genetically, often show deficits in the formation of organized place fields.
Finally, although visual cue is very important, it is also observed that place cell firing persist even in the dark, which implys that other sonses (or proprioception) may contribute to the firing as well.
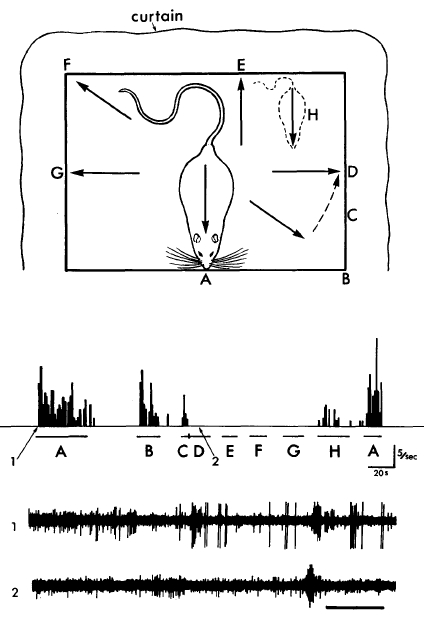

A schematic of an early place cell experiment and some data demonstrating the direction specific nature of this place cell's response. (From: O'Keefe and Dostrovsky)
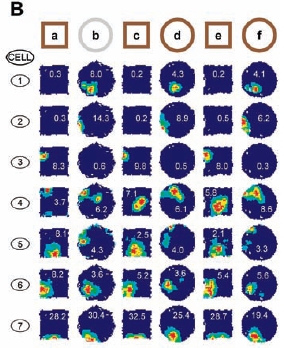

The evolution of the place fields of 7 different cells as a mouse is repeatedly exposed to two different environments. (From: Wills, O'Keefe, et al.)
Other Neurons with Spatial Firing Propertis
Grid cells
Grid cells are first discovered in 2005 by Hafting et al. A grid cell is a type of neuron that fires strongly when the animal is in a particular location in the environment. It is found in the entorhinal cortex and it is hypothesized that these grid cells could connect and produce a mental map of this particular environment. Although grid cells are found in all layers of entorhinal cortex, layer II cells are more grid-like and they are independent of the head direction of the animals.
Hafting T, Fyhn M, Molden S, Moser MB and Moser EI. Microstructure of a spatial map in the entorhinal cortex. Nature 2005 436, 801-6
Head direction cells
Head direction cells are related to place cell system. They are neurons that fire when the animal's head points in a specific direction within an environment. The firing is independent of the body position relative to the head. Unlike place cells, head direction cells do not require a functional hippocampus. They are found in many regions, including post-subiculum, retrosplenial cortex, anterior thalamis nucleus, lteral dorsal thalamic nucleus, lateral mammillary nucleus, dorsal tegmental nuclues, and striatum.
The anatomical and computational basis of rat HD signal. Sharp et al. 2002
References
Ekstrom A, Kahana M, Caplan J, Fields T, Isham E, Newman E, Fried I. Cellular networks underlying human spatial navigation. Nature. 2003 Vol 425, 184-188.
Wills TJ, Lever C, Cacucci F, Burgess N, O'Keefe J. Attractor dynamics in the hippocampal representation of the local environment.Science. 2005 May 6;308(5723):873-6.
Independent rate and temporal coding in hippocampal pyramidal cells. Nature. 2003 Oct 23;425(6960):828-32.
Lever C, Wills T, Cacucci F, Burgess N, O'Keefe J. Long-term plasticity in hippocampal place-cell representation of environmental geometry. Nature. 2002 Mar 7;416(6876):90-4.
The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971 Nov;34(1):171-5.
Recommended reading: Bures J, Fenton AA, Kaminsky Y, Zinyuk L. Place cells and place navigation. Proc Natl Acad Sci U S A. 1997 Jan 7;94(1):343-50.
Recent updates to the site:
List of abbreviations:
- N
- This edit created a new page (also see list of new pages)
- m
- This is a minor edit
- b
- This edit was performed by a bot
- (±123)
- The page size changed by this number of bytes
11 June 2026
| 02:22 | Labs diffhist +52 Mohammad H. Dezfulian talk contribs (→North America) | ||||
|
|
02:14 | User:Mohammad H. Dezfulian 7 changes history −4,949 [Mohammad H. Dezfulian (7×)] | |||
|
|
02:14 (cur | prev) +416 Mohammad H. Dezfulian talk contribs (→Contact Info) | ||||
|
|
02:12 (cur | prev) −269 Mohammad H. Dezfulian talk contribs (→Fellowships) | ||||
|
|
02:12 (cur | prev) −5,151 Mohammad H. Dezfulian talk contribs (→Conference Presentations & Proceedings) | ||||
|
|
02:12 (cur | prev) −880 Mohammad H. Dezfulian talk contribs (→Academic Honors) | ||||
|
|
02:11 (cur | prev) −409 Mohammad H. Dezfulian talk contribs (→Funding) | ||||
|
|
02:11 (cur | prev) +1,357 Mohammad H. Dezfulian talk contribs (→Peer Reviewed) | ||||
|
|
02:07 (cur | prev) −13 Mohammad H. Dezfulian talk contribs (→Contact Info) | ||||
