BMCB625:Helicases

Bonus Materials
Review PCNA, the Maestro of the Replication Fork [[1]] A recent addition to Cell that looks at a critical component of the replication fork. A fairly good review. --Chris 11:39, 18 May 2007 (EDT)
(Homework) Questions
Chris
“Previous work has demonstrated that…” E1 binds a site-specific dsDNA and then ssDNA followed by its “known helicase activity.” (1) How does one demonstrate this experimentally (see paper ref. 6, 19-21), (2) what are the barriers to overcome energetically seeing as how the clamp appears relatively stable once bound to DNA (i.e., is there a particular “location” where the protein subunits “come apart” after being pre-formed or are they assembled subunit-by-subunit at the specific binding site? Either way, is there additional factors that help assemble this complex or can it freely form in solution on its own?
Mahta
Q1. Separation of the duplex into ssDNA by the helicase via the proposed model utilizes one ATP for each base pair translocated through the center of the hexamer. Is this not an inefficient use of energy?
A1. It can both be thought of as an inefficient and efficient use of energy. In terms of it being inefficient - this system allows for one nucleotide to be pushed through the helicase at the expense of one molecule of ATP. That is a huge input of energy for this process. However, if one considers the importance of this process (the importance of accurately carrying out separation of strands for replication, transcription, etc) the energy expenditure does not seem by any means unnecessary. It can also be a matter of making sure these processes are carried out with great accuracy and a very high speed.
Q2. Is it clear whether the helicase hexamer is "pushing" the ssDNA through its center core or is the helicase itself translocating along the DNA strand? (See Fig S-5) The basis for this question lies in their claim that in vivo there are two hexamers - in one, all six subunits can bind DNA whereas in the second one only five of six subunits can bind DNA - does this favor "DNA pushing through" or the helicase translocating itself on the DNA?
Chayne
In Enemark & Joshua-Tor, they propose the "escort" model as opposed to the "bucket brigade". Since this is just conjecture based upon observations from structure (though a pretty good one), how could this model be more thoroughly tested?
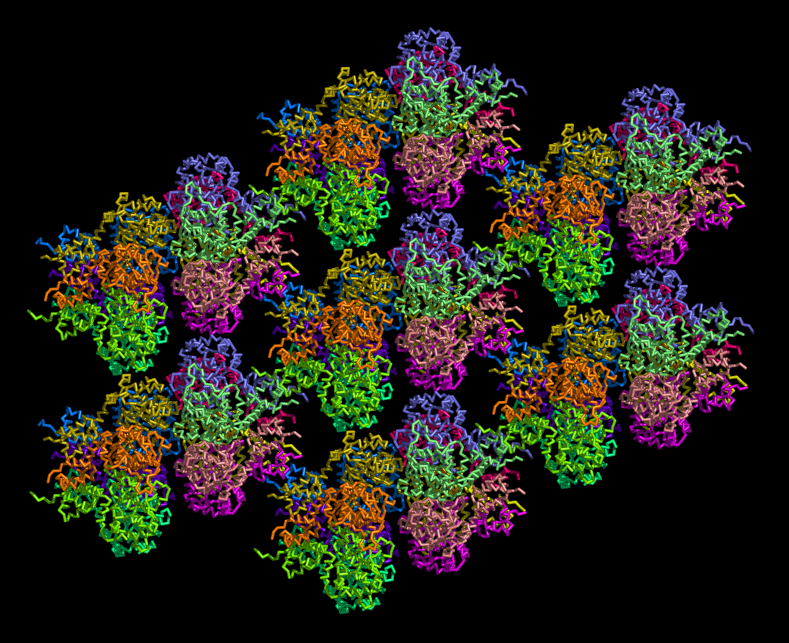
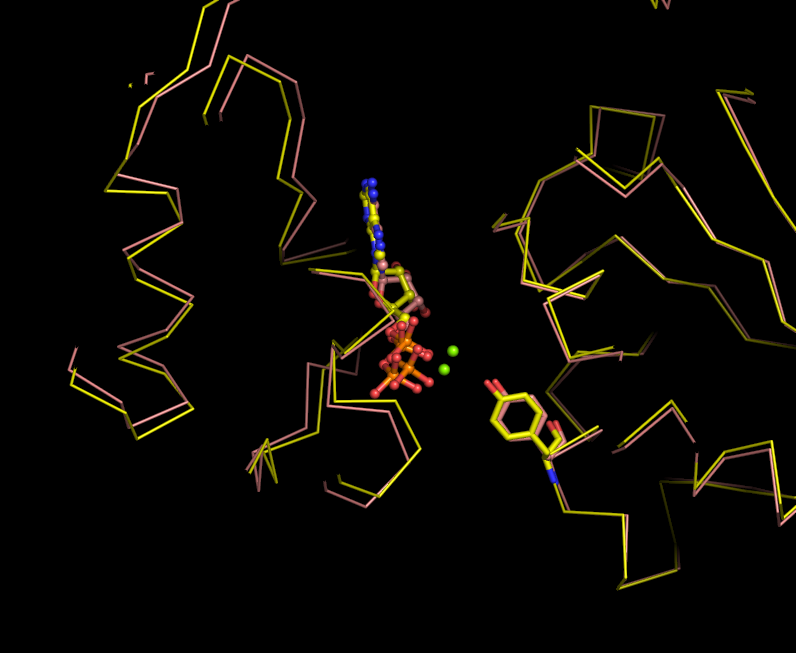
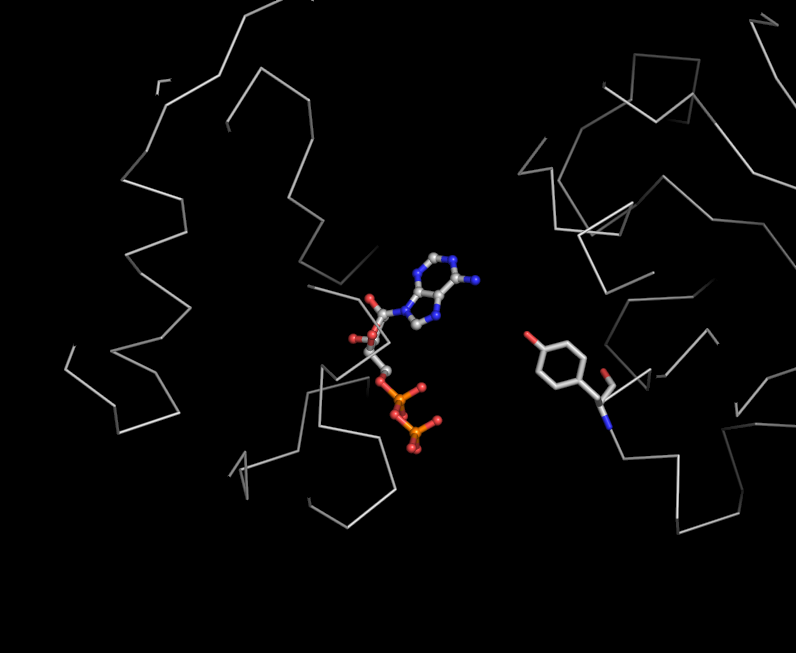
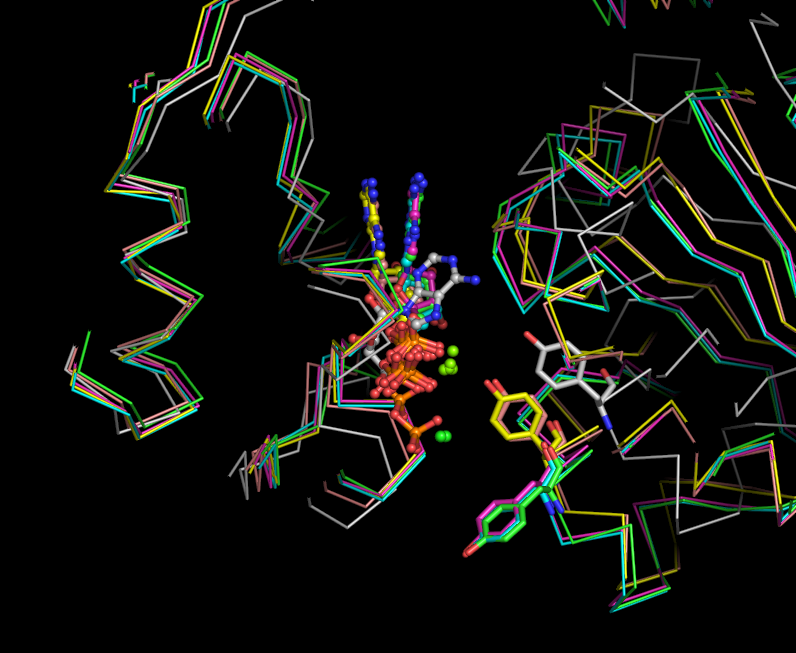
some figures I made from pdb 2GXA (not really related to above question)
Media:Fig2-2GXA-loop-superpose.pdf
Media:Fig3-ADP_bs-lsqDE-ADPlike.png
Media:Fig4-ADP_bs-lsqABC-ATPlike.png
Media:Fig5-ADP_bs-lsqF-APOlike.png
Media:Fig7-ADP_bs-lsqall-view2.png
Jeremy
Q1. Does the fact that helicase subunits are stabely bound by ADP in the crystal hint at something mechanistically important?
A1. The fact the structure is stable when bound by ADP suggests a stable intermediate, mechanistically. In other words, an unloading step, where ADP is removed so that ATP may enter may be required for helicase function.
Q2. The structure reveals several polar contacts between the helicase and DNA, but highly non-polar residues are within VDW distance of the DNA. However counter-intuitive, why might this be important?
A2. Polar contacts are necessary to obtain a grip on DNA, the non-polar, or greasy contacts however, may provide speed for rapid helicase function. There may be a balance between polar and non-polar contacts to allow rapid unwinding, where the DNA slides across the greasy surfaces.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}