Centromeres: Controllers of Inheritance
(You can also read about our research at the HHMI website)
We study the fundamental biology of genetic inheritance, and aim to manipulate it for practical benefit. Centromeres control chromosome segregation during cell division, because they are the loci at which chromosomes attach to spindle microtubules via the kinetochore protein complex. Centromere DNA in most plants and animals consists of megabases of simple tandem repeats. These sequences can be dispensable for centromere function. Instead, the centromere is epigenetically specified by CENH3, a centromere-specific histone H3 variant that replaces conventional H3 in centromeric nucleosomes, and is essential for recruiting other kinetochore proteins.
The plant Arabidopsis thaliana is ideal for studying chromosome segregation, because it combines facile genetics and cytology with large centromeres that are similar to those vertebrate cells (by contrast, laboratory yeasts have very small centromeres).
We have discovered that centromere differences between two parents can cause massive chromosome segregation errors when their genomes meet in the fertilized zygote. When Arabidopsis plants expressing altered CENH3 proteins are crossed to wild type, chromosomes from the mutant parent can be completely eliminated, yielding haploid progeny. Major research projects in the lab are listed below.
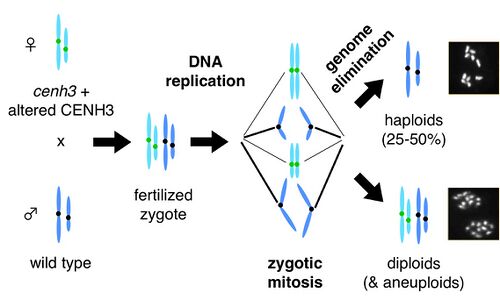
1) Mechanism of genome elimination
We are using genetic and cytological methods to investigate chromosome missegregation caused by parental centromere differences. Crosses between cenh3 mutants and wild-type feature a high amount of seed abortion, and produce a mixture of diploid, aneuploid and haploid progeny. We are working to understand the mechanistic basis of these observations.
2) Centromere evolution
Despite the essential nature of centromeres, their DNA sequence and the CENH3 protein evolve rapidly. To study the functional consequences of rapid centromere evolution, we are manipulating CENH3 and centromere DNA in Arabidopsis and in other plants. In collaboration with Ian Korf, we are also using comparative genomics to study how centromere DNA evolution has been constrained by functional demands.
3) Chromosome engineering to create new plant breeding technologies
Haploid plants that are converted back into diploids can greatly accelerate plant breeding. Such “doubled haploids” produce instant homozygous lines from heterozygous F1s, a process that normally takes 8-10 generations of inbreeding. As described above, we have discovered a simple method for producing haploid plants through seed by manipulating CENH3. Haploids are easily converted to diploids, so Arabidopsis geneticists can produce large recombinant populations in a single step, or rapidly generate multiple mutants (e.g. 1 out of 256 haploid progeny will contain eight unlinked mutations if one starts with a octuple heterozygote). A detailed protocol for generating Arabidopsis haploids is available at http://tinyurl.com/ArabidopsisHaploidProtocol
Our method has key advantages over current procedures that often require tissue culture and are limited to specific species or genotypes. As CENH3 is found in all eukaryotes, the procedure should theoretically work in any plant species. To learn more about our technology, please see this website.
We are also interested in developing new plant breeding methods through chromosome engineering. Hybrid vigour is the basis of many high-yielding agricultural varieties, but it is impossible to propagate a heterozygous genotype through sexual reproduction. In collaboration with Imran Siddiqi and Raphael Mercier's labs, we have recently shown that hybrid Arabidopsis can be crossed to a cenh3 mutant to create clonal progeny that preserve their heterozygous genotype (see Publications for details).
| 
