Protein domains exhibit a wide variety of physicochemical properties and represent a rich source for delivery of the specific function to a defined position on DNA origami. In order to demonstrate the proof of the concept for spatial addressing of protein functions to a DNA origami, we considered using restriction enzymes, proteases, biosynthetic enzymes, proteins with optical properties, motor proteins etc.
Finally we decided to explore the performance of BRET - bioluminescence energy transfer on the DNA origami. BRET is a biophysical phenomenon in which energy is non-radiatively transferred from the donor (Renilla luciferase in our case) to the acceptor (mCitrine variant of the yellow fluorescent protein) when both molecules are in close proximity (within 10 - 100 Å range). Renilla luciferase (RLuc) catalyzes the reaction of oxidative decarboxylation converting its substrate coelenterazine to coelenteramide by the simultaneous emission of light with a maximum emission peak around 475 nm. The energy of photons is high enough to excite the yellow fluorescent protein (in case it is bound within the required distance from the Renilla luciferase) leading to fluorescence with an emission maximum of 529 nm. Upon successful implementation of binding of BRET pair to DNA origami, the system could be expanded to four partners by splitting each of Renilla luciferase and mCitrine into 2 segments, where BRET would depend on the proximity of four sites (two pairs). Because of the distance-dependent signal efficiency, BRET could be employed as a nanomolecular ruler or as a logical operator for <a
href="http://openwetware.org/wiki/Biomod/2011/Slovenia/BioNanoWizards/appbiosensors">biosensors</a> as outlined in the Applications section.
<img
style="padding: 0pt 0pt 5px; width: 890px; height: 238px;" alt="" title=""src="http://openwetware.org/images/9/9a/BRET_final.png"> |
| Figure 18: Functionalized ZFPs decorating DNA origami plane. (a) Molecular models of BRET triple fusion proteins, namely MBP-RLuc-2C7 and MBP-mCitrine-AZPA4, are presented. BRET is observed after Renilla luciferase catalyses oxidative decarboxylation of its substrate coelenterazine to coelenteramide which in terms emits light at ~ 475 nm and excites mCitrine when in close proximity (1 - 10 nm) (b) DNA origami plane decorated with BRET partner fusions. |
We first prepared constructs for BRET protein chimeras in the form of RLuc-2C7 and YFP-AZPA4. Unfortunately, it turned out the two proteins were expressed exclusively in the insoluble fraction of the cell lysate (Figure 19), which means that we would need an efficient refolding procedure to prepare functional proteins for binding to DNA origami. We tried several isolation and refolding procedures with little success.
Next, we tried to optimize the recombinant protein production conditions in order to purify the produced protein fusions in the native form. A well-known strategy for enhancing solubility and proper folding of larger protein fusions during their production is to lower the temperature as well as the concentration of the inducer IPTG allowing the protein to fold correctly and decrease the formation of protein aggregates in vivo. We achieved partial expression of soluble YFP-AZPA4 (Figure 20), with no improvement on solubilization of RLUC-2C7 fusion.
<img
style="width: 276px; height: 374px;" alt=""src="http://openwetware.org/images/1/1f/InsolubleBRETfusionsWBproduction.png"> |
<img alt="" src="http://openwetware.org/images/6/66/MBPYFPAZPA4alteredcond.png"> |
| Figure
19. ZFP-luciferase and ZFP-YFP fusions are produced in bacteria in an insoluble form. Figure shows Western blot after production of fusion proteins. Arrows to the left indicate the position of bands representing BRET fusions without MBP solubility tag present in an insoluble form (IBs - inclusion bodies). Grey arrow depicts the position of YFP-AZPA4 (Mr of 48 kDa) and black arrow the position of RLuc-2C7 (Mr of 59 kDa). Production conditions: 2x YT medium supplemented with 0.1 mM ZnCl2 and 100 mg/L antibiotic kanamycin / 37 °C / 180 rpm / ~7 h induction with 2 mM IPTG. |
Figure
20. Production of low amount of soluble YFP-AZPA4 fusion by modification of production conditions. Western blot after production of YFP-AZPA4 with altered fermentation conditions (LB medium supplemented with 0.5 mM ZnCl2 and 100 mg/L antibiotic kanamycin / 24 °C / 160 rpm / ~24 h induction with 0.5 mM IPTG). This made YFP-AZPA4 protein (Mr of 48 kDa) observable in the soluble form (depicted by the arrow to the right), but not to sufficient extent to isolate it. |
Since we have already observed the beneficial effect of enhanced solubility by attaching MBP and/or GST domains to the N-terminal end of zinc finger proteins (section <a
href="http://openwetware.org/wiki/Biomod/2011/Slovenia/BioNanoWizards/resultssolublezfp">Soluble
ZFPs</a>), MBP domain was added to the N-termini of both BRET fusion proteins, creating triple fusion proteins in the following form: MBP-RLuc-2C7 and MBP-YFP-AZPA4, aiming to increase their solubility and facilitate the isolation of functional proteins under native conditions.
We anticipated that decreasing the temperature during protein production would lead to further increase of final protein titers but ultimately we observed the solubility of both triple chimeras was comparable at 30 °C and 37 °C as can be inferred from the results below. We used the following conditions for the production: 2x YT medium supplemented with 10 g/L glucose, 100 mg/L antibiotic kanamycin and 0.5 mM ZnCl2 / 30 °C or 37 °C / 160 rpm / ~5 or 7 h induction with 1 mM IPTG.
<img
style="margin-bottom: 10px; width: 593px; height: 287px;" alt=""src="http://openwetware.org/images/0/01/BRETfusionswithsolubilityTagproductionCBB.png"> |
| Figure 21. ZFP-BRET-MBP fusions are produced in soluble form. Bacterial lysate fractions after production of BRET fusions with MBP domain were analyzed with Coomassie Brilliant Blue stained SDS PAGE analysis. Grey arrow to the right indicates the position of MBP-YFP-AZPA4 (Mr of 90 kDa) and black arrow the position of MBP-RLuc-2C7 (Mr of 101 kDa). Addition of MBP solubility tag promoted the presence of both protein chimeras in supernatant (SN). Both proteins were partially present in inclusion bodies (IBs) which was the case with all other ZFP chimeras as well. |
<img
style="margin-bottom: 10px; width: 558px; height: 287px;" alt=""src="http://openwetware.org/images/0/0e/BRETfusionswithsolubilitytagproductionWB.png"> |
| Figure 22. ZFP-BRET-MBP fusions are produced in soluble form. Western Blot after production of BRET fusions with MBP tag. Grey arrow to the right indicates the expected position of MBP-YFP-AZPA4 (Mr of 90 kDa) and black arrow the position of MBP-RLuc-2C7 (Mr of 101 kDa). |
<img
style="margin-right: 20px; width: 200px; height: 306px;" alt=""src="http://openwetware.org/images/6/63/BRETfusionsisolationCBB.png"> |
Figure 23. Isolation of BRET fusions with MBP tag using chelating chromatography. Coomassie Brilliant Blue (CBB) stained SDS PAGE analysis of isolated fractions. Isolation of both BRET triple fusion proteins (MBP-YFP-AZPA4, Mr of 90 kDa and MBP-RLuc-2C7, Mr of 101 kDa) resulted in a single protein band observed after staining the SDS-PAGE gel with CBB. The yield of MBP-YFP-AZPA4 was approximately 10-times higher compared to MBP-RLuc-2C7. |
After protein isolation functional assays were performed to analyze the effect of added protein domains at both N- and C- terminus of yellow fluorescent protein (mCitrine) and Renilla luciferase (RLuc). We determined the fluorescence of YFP fusions and decarboxylation enzymatic activity on the luciferase's substrate coelenterazine leading to light emission in case of luciferase.
Characterization
of MBP-YFP-AZPA4
<img
style="margin-right: 20px; width: 450px; height: 223px;" alt=""src="http://openwetware.org/images/7/72/MBPYFPAZPA4fluorescentspectrum.png"> |
Figure 24. Fluorescence emission spectrum of MBP-YFP-AZPA4 fusion. The fluorescence emission spectrum of MBP-YFP-AZPA4 chimera was acquired on a PerkinElmer LS55 Luminescence Spectrometer. Protein sample was excited at 485 nm and emission spectra were obtained in the 500 to 600 nm window with a scanning speed of 100 nm/min. Red line represents blank control (lysis buffer), green line is a negative control (2C7-MBP-6F6 supernatant) and blue line shows the fluorescence emission spectrum of MBP-YFP-AZPA4 chimera. |
<img
style="margin-bottom: 10px; width: 750px; height: 254px;" alt=""src="http://openwetware.org/images/2/23/MBPYFPAZPA4visual.png"> |
| Figure 25. Visual characterization of the isolated MBP-YFP-AZPA4 chimera under UV light. Left: centrifuge tubes with isolated MBP-YFPA-AZPA4 (50, 100 and 250 mM imidazole elution fractions) were illuminated with UV light. Negative control (isolated AZPA4-MBP-6F6) was added to the left of the three fluorescent samples to demonstrate the difference in optical properties of isolated proteins. Right: 200 μl of 100mM imidazole elution fraction of MBP-YFP-AZPA4 was pipetted into Corning Costar 96-well white microtiter plate with transparent bottom to yield a pattern of "YFP". The plate was analyzed in DNA Bio-Imaging Systems box by illumination with the UV light. |
Characterization
of MBP-RLuc-2C7
<img
style="margin-right: 20px; width: 400px; height: 307px;" alt=""src="http://openwetware.org/images/2/21/MBPRLuc2C7biolum.png"> |
Figure 26. MBP-RLuc-2C7 ZFP chimera exhibits bioluminsecence. 40 μl of MBP-RLuc-2C7 supernatant (non-diluted, diluted 2 or 5-fold) was transferred to Corning Costar 96-well white microtiter plate. The bioluminescence assay was performed on a Berthold's ORION II Microplate Luminometer. |
<img
style="margin-right: 20px; width: 780px; height: 255px;" alt=""src="http://openwetware.org/images/8/83/MBPRLuc2C7visual.png"> |
| Figure 27. Visual characterization of the activity of MBP-RLuc-2C7 chimera.
Left: 100 μl of MBP-RLuc-2C7 supernatant was transferred to Perkin Elmer OptiPlateTM 96-well white plate in a pattern giving rise to the word "Luc". Right: 100 μl of the coelenterazine h substrate was added to the protein sample prepared in the same way as for the bioluminescence assay. Luminescence was detected in Syngene G:Box for 5 sec. |
BRET effect to detect neighboring binding sites on DNA target
<img
style="margin-right: 20px; width: 780px; height: 133px;" alt=""src="http://openwetware.org/images/3/32/BRETDNAtarget.png"> |
| Figure 28: Sequence of the DNA target designed for BRET experiment.
43 bp DNA target used to measure the BRET effect was designed as shown above. 2C7 binding sequence is colored blue and AZPA4 binding site in green. Spacer of 2 bp was selected to separate the binding sites and a clamp of the same length at both 5' and 3' end. |
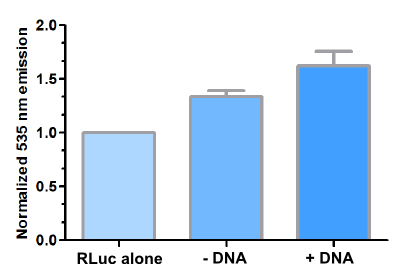
<img
style="margin-right: 20px; width: 400px; height: 260px;" alt=""src="http://openwetware.org/images/b/b8/BRETgraphDNA.png"> |
Figure 29: BRET resulting from the addition of DNA target sequence to chimeric ZFP fusions. mCitrine emission was normalized to the 535 nm emission of the MBP-RLuc-2C7. Since the emission spectra of RLuc and mCitrine overlap, 535 nm emission is observed for 100 nM MBP-RLuc-2C7 as well. Addition of 50 nM DNA target increased mCitrine emission indicating proximal binding of chimeric proteins to the DNA target. |
We demonstrated the design, production, purification and characterization of the two protein elements required for the reconstitution of the BRET sensor on a DNA origami. Initial experiments support the functional BRET, however the overlapping Renilla luciferase and mCitrine emission spectra suggest that selection of other BRET partners might improve the signal to background ratio, e.g. replace the Rluc-YFP pair with RLuc8-GFP2 BRET pair as a reasonable choice (De, 2007). Sensitivity of the BRET experiment on DNA origami could be improved by binding several neighboring BRET pairs on a single DNA origami rectangle.
- De A, Loening AM, Gambhir SS. (2007) An Improved Bioluminescence Resonance Energy Transfer Strategy for Imaging Intracellular Events in Single Cells and Living Subjects. Cancer Research, 67: 7175-7183.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
}){kind=link}
}){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}