IGEM:IMPERIAL/2008/Bioprinter/Subteam 2
Hey guys - ran into this online and thought it might be useful for seeding light-sensor searches. It's the PhD thesis of a Dutch student: Towards Unveiling the Photoactive Yellow Proteins: characterization of a halophilic member and a proteomic approach to study light responses ~ Tom Adie 15:45, 21 July 2008 (UTC)
Tasks
- Summarise pathways of the different types of photoreceptors found in bacteria. Please take a look at this review paper: Photosensing in chemotrophic, non-phototrophic bacteria: let there be light sensing too [1]
- Find links between photoreceptor pathways and well-studied pathways in B.Subtilis
- Obtain reaction kinetics and determine responsiveness of photoreceptor molecules and associated molecules.
Findings
- Main classes of photoreceptors
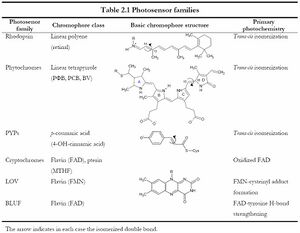
Main classes of photoreceptors (adapted from S.Memmi)

The most commonly studied photoreceptors include: rhodopsin (Archaea), phytochromes (cyanobacteria), photoactive yellow proteins, LOV and BLUF. Most of these function through the binding of a photon which causes a conformation change, often in the form of photoisomerisation. Others exhibit oxidation or H-bond strenghtening upon photon binding.
This review paper gives a brief outline of the YtvA-σB activation pathway along with a list of other photoreceptors found in bacteria which you will find helpful in your research. YtvA is naturally present in B.Subtilis and contains a blue light sensing LOV domain. Note that proper care must be taken when exposing B.Subtilis to light, since the pathway is activated under very low intensities of blue light.
This paper talks about the different type of genes regulated by σB which is produced as part of a stress response. The main areas affected include DNA protection, envelope formation, regulation of influx and efflux, carbon metabolism and macromolecule turnover. There is little indication though, of sporolation (although it is suggested to be regulated as well) and motility.
- Phytochrome / Engineering bacteria to see light [3]
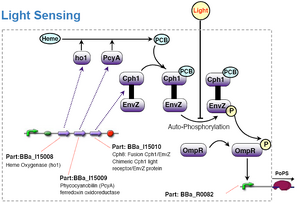
Phytochrome red light sensor

Phytochromes possess a bilin prosthetic group whose activity allows it to photoisomerise into two states: red-light absorbing (Pr) and a far-red-light absorbing (Pfr). All phytobilins are derived from haem, which is converted to biliverdin IXα by haem oxigenase HO1 and then reduced by a bilin reductase PcyA. This paper describes the genetic engineering of phytochrome biosynthesis in E.Coli [4].
In 2005 a team successfully introduced phytochrome (from cyanobacteria) which is a red-light receptor, into E.Coli. The team based this on a well studied system in E.Coli. By creating a Cph1(phytochrome)-EnvZ chimera, they essentially constructed a red-light input - reporter output system. They have also submitted two parts BBa_I15008, BBa_I15009 into the MIT registry. Parallels can be drawn by introducing a naturally occuring red-light photoreceptor into B.Subtilis, however, it should be noted that the phytochrome is a membrane-bound extracellular photoreceptor. Hence, even though it might have worked with E.Coli, it might not neccessarily work with B.Subtilis. The pathway is as shown:
- AppA

Appa Pathway (Image from Masuda et al)

Within the AppA system the effect of light on gene expression is mediated through the repressor PpsR4. From the diagram it can be seen that the system is only light sensitive in anaerobic conditions, in these conditions exposure to UV (blue light) causes the disassociation of PpsR4 from AppA and allow the PpsR4 to bind DNA and cause repression. [5]. But when this Rhodobacter system has been reconstructed exogenously in E.coli the system fails to respond to blue light [6]
- Rhodopsin
There exists four functionally different rhodopsins: bacteriorhodopsin (BR ~568nm), halorhodopsin (HR ~578nm), sensory rhodopsin I (SRI ~587nm) and sensory rhodopsin II (SRII ~487nm) and all of them have a seven helix membrane spanning structure. Rhodopsins are commonly found in archea, in particular, the most commonly studied Halobacterium salinarum. Bacteriorhodopsin is a proton pump, halorhodopsin a chloride pump, while the last two control bacterial chemotaxis away from damaging UV light. SRII and HtrII are both membrane spanning proteins. When SRII absorbs blue light, conformational changes are transferred to HtrII which then regulates cell motility.
In Archaea, SRI and SRII mediate phototactic responses. When oxygen and respiratory substrates are abundant, SRII is synthesised, allowing the organism to evade any form of photo-oxidative damage. When there is a drop in oxygen, SRII synthesis is inhibited and SRI synthesis is activated and together with BR and HR, generate a proton motive force for ATP synthesis. SRI stimulates a phototactic response towards areas illuminated by orange light. Due to an intermediate of SRI which absorbs near-UV light and elicits a repellent response, phototaxis towards orange light only occurs in the absence of near-UV light.
- Photoactive Yellow Protein

PYP Cycle(Adapted from S. Memmi)

PYP from H.halophilia is the best characterised Photoactive Yellow Protein. It is a small sized protein (125 AAs), is water-soluble and is stable along a broad pH range. The photochemical properties of PYP resembles sensory rhodopsin and it absorbs blue light at ~446nm. Upon illumination by blue light, PYP undergoes a series of cyclic reactions as shown in the figure. Other factors which regulate the performance of PYP include temperature, pH, hydrophobicity and viscosity. Although it may seem that PYP has provided us a perfect solution to the problem of membranous photoreceptors, the way PYP regulates gene expression in bacteria remains unknown.
References:
- Romero G. Inositolglycans and cellular signalling. Cell Biol Int Rep. 1991 Sep;15(9):827-52. DOI:10.1016/0309-1651(91)90036-i |
- Price CW, Fawcett P, Cérémonie H, Su N, Murphy CK, and Youngman P. Genome-wide analysis of the general stress response in Bacillus subtilis. Mol Microbiol. 2001 Aug;41(4):757-74. DOI:10.1046/j.1365-2958.2001.02534.x |
- Levskaya A, Chevalier AA, Tabor JJ, Simpson ZB, Lavery LA, Levy M, Davidson EA, Scouras A, Ellington AD, Marcotte EM, and Voigt CA. Synthetic biology: engineering Escherichia coli to see light. Nature. 2005 Nov 24;438(7067):441-2. DOI:10.1038/nature04405 |
- Gambetta GA and Lagarias JC. Genetic engineering of phytochrome biosynthesis in bacteria. Proc Natl Acad Sci U S A. 2001 Sep 11;98(19):10566-71. DOI:10.1073/pnas.191375198 |
- Masuda S and Bauer CE. AppA is a blue light photoreceptor that antirepresses photosynthesis gene expression in Rhodobacter sphaeroides. Cell. 2002 Sep 6;110(5):613-23. DOI:10.1016/s0092-8674(02)00876-0 |
- Jäger A, Braatsch S, Haberzettl K, Metz S, Osterloh L, Han Y, and Klug G. The AppA and PpsR proteins from Rhodobacter sphaeroides can establish a redox-dependent signal chain but fail to transmit blue-light signals in other bacteria. J Bacteriol. 2007 Mar;189(6):2274-82. DOI:10.1128/JB.01699-06 |
Useful Reviews:
<biblio>
- review1 pmid=16228399
- review2 pmid=15170486
- review3 pmid=9297461
- review4 pmid=15170480
