CH391L/S2013 Taylor Pursell Feb 27 2013
Identification of Early Replicating Fragile Sites that Contribute to Genome Instability. Barlow JH, Faryabi RB, Callén E, Wong N, Malhowski A, Chen HT, Gutierrez-Cruz G, Sun HW, McKinnon P, Wright G, Casellas R, Robbiani DF, Staudt L, Fernandez-Capetillo O, Nussenzweig A. Cell. 2013 Jan 22. pii: S0092-8674(13)00008-1.
Background
B-Lymphocytes
B lymphocytes, or B-cells, originate in the bone marrow. The B-cells are then circulated in the blood and become activated through interactions with an antigen and/or T-cells. In this case we will focus on T-cell dependent activation where B-cells undergo terminal maturation after becoming activated through interaction with an antigen and a T-cell. Once activated, large B-cells undergo rapid proliferation and genomic rearrangements important for expansion and specification of the antibody repertoire [1,4,5]. These genome changes come in two classes:
- Somatic Hypermutationi(SHM) in which point mutations, or more rarely deletion or insertions, are induced in the variable region of immunoglobulin (Ig) genes and their surrounding regions.
- Class Switch Recombination (CSR) where one constant region of Ig is deleted and replaced with another[1,5].
Activation-Induced Cytidine Deaminase and DNA Damage
An enzyme called activation-induced cytidine deaminase (AID) is required for SHM and CSR to occur [5,6]. By deaminating cytosine residues on ssDNA that are exposed during Ig transcription, the enzyme turns the cytosine residue into a uridine. The mismatched uridine is glycosylated and removed by uracil DNA glycosylase (UNG) allowing the apyrimidic endonuclease (APE) to generate the DNA breaks needed for SHM and CSR to occur [5].
AID has been shown to act in the presence of a particular motif: DGYW (D= any bas but C ; G= guainine; Y= pyrimidine ; W= adenosine or thymidine) [6]. Although this is its primary function, it is not uncommon for it to act on other DNA segments containing motif including oncogenes like c-Myc [1].
Replication and Fragile Sites
The eukaryotic genome is divide into regions which are programmed to replicate at different times during S-phase of the cell cycle. There are two groups of replication origins: early firing origins and late origins. Origins near active genes, or genes whose products are important to the cell, fire in the first half of S-phase in a process called early replication. In contrast, origins near inactive genes fire during the second half of S-phase and are associated with late replication[2].
Chromosomal fragile sites are specific stretched of genetic material that are prone to gaps and breaks following replication leading to chromosome instability; these fragile sites are put into two classes: rare and common fragile sites. Rare fragile sites are those found in only a few individuals and are usually associated with genetic disorders while common fragile sites (CFS) are found in all individuals [3]. Common fragile sites are can be associated with large genes, long stretches of AT rich regions, and stalled DNA replication [1,3]. Common fragile sites are associated with late replicating origins [1,2,3].
Introduction
In studies preformed in yeast, origins at which replication fork collaspe occurs in response to stalled replication have been identified [7]. These origins do not line up with "replication slow zones" or the yeast equivalent of CFS found at late replication origins [8,9] suggesting there is another class of fragile sites in yeast.
The researchers in this study identify similar early replicating origins at which DNA lesions are found in B-cells; they termed these sites early replicating fragile sites (ERFSs) [1].
Methods
ChIP and ChIP-Seq

Mouse splenic B-cell samples were stimulated with LSP/IL4 to cause the cells to synchronously enter cell cycle. After 22 hours, cells should be in S phase (monitored by FACs). Cells were then treated with 10 mM hydroxyurea (HU) which arrests cells at G1/S phase and then cells were harvested. Cells were crosslinked in order to ensure any protein bound to DNA would remain bound. Cells are sonicated in order to break up cells and to fragment DNA. Sonicated chromatin samples are then incubated with an antibody specific to the protein of interest (e.g. RPA, BRCA1, etc.) and magnetic breads specific to the constant region of the antibody used. These magnetic beads are used to pull out or precipitate the anitbody, protein, and bound sequence of DNA. The protein and DNA can be separated from the bead and prepared for sequencing. For schematic see figure to the right from the manufacture's website (http://www.illumina.com/technology/chip_seq_assay.ilmn).
Once genetic material is harvested, it was prepared for micro sequencing using Illumina’s protocol for sample preparation and was analyzed using the Genome Analyzer IIx.

Fluorescent in-situ hybridization (FISH) analysis
Fluorescent probes that bind to complementary sequences on chromosomes are used to visualize abnormalities. Fluorescent microscopy can be used to find where the probe is bound. In this paper, blue is DAPI-stained DNA, green is the insert in the BAC (either ERFSs or CFSs) and red is telomeric DNA.
Results
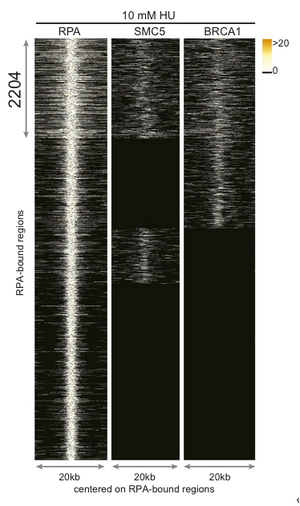
First researchers needed to map early replication origins in an activated B-cell genome. To do this they performing ChIP with anti-replication protein A (RPA). They harvested fresh mouse splenic B-cells, stimulated them with LPS/IL4 to enter cell cycle. They then inhibited replication in G1/S phase using hydroxyurea (HU) allowing RPA binds to ssDNA at stalled forks. In addition to RPA, the researchers looked for the localization of recombination proteins BRCA1 and SMC5 to regions also occupied by RPA in order to identify regions in which the fork was not only stalled but had collapsed and required homologous recombination repair. From this ChIP data, they were able to identify 2204 regions were RPA, BRCA1, and SMC5 were all localized (Figure 1D).
Upon further characterization of the 2204 regions, it was found that they contained shared common elements such as highly repetitive sequences (e.g. LINE L2, SINE, DNA transposon, and tRNA elements) which are known fork barriers and are significantly more GC rich in contrast to the rest of the genome. This also distinguishes them as their own class from CFSs which are AT rich regions [1,3].
Because most (80%) of ERFSs are within 300 kb of one another, researchers grouped the ERFSs into regions they call ERFS hot spots. They then narrowed the hotspots down to those with a ChIP footprint larger then 10kb to give 619 hotspots to hot spots scattered throughout the genome (Figure 2B, below). The top 15 hotspots with the highest RPA/BRAC1/SMC5-binding strength were analyzed further to reveal that 8 out of 15 rearranged in B-cell lymphoma (see table 2C) [1].

Significance/Applications
References
- Barlow, J. H. et al. Identification of Early Replicating Fragile Sites that Contribute to Genome Instability. Cell 152:1-13 (2013)
- Goren A, Cedar H. Replicating by the clock. Nature Reviews Molecular Cell Biology In Press 4: 25-32 (2003)
- Durkin, S. G. & Glover, T. W. Chromosome Fraglie Sites. Annual Reviews Genetic 41: 169-192 (2007)
- Parham, Peter. The Immune System. Garland Science, 2. (2005)
- Durandy, A. Activation-induced cytidine deaminase: a dual role in class-switch recombination and somatic hypermutation. Eur. J. Immunol 33: 2069-2073 (2003)
- Kenter, A. L. & Bhattacharya, P. AID: a very old motif newly recognized. Nature Immunology 5: 1203-04 (2004)
