Biomod/2013/NanoUANL/Enzyme
<html> <head> <link href='css/left_menu.css' rel='stylesheet' type='text/css'> </head>
<style> .main_cont { float:left; width:150px; background-color:#4d7986; padding:10px; } .menu_top_bg { width:150px; background:url(http://www.cssblog.es/images/menu_top_bg.gif) repeat-x; height:22px; padding-top:8px; font-family:Verdana, Arial, Helvetica, sans-serif; font-size:12px; color:#FFFFFF; font-weight:bold; text-align:center; margin-bottom:1px; } .sub_menu ul { padding:0px; margin:0px; } .sub_menu ul li { font-family:Arial, Helvetica, sans-serif; font-size:11px; color:#FFFFFF; line-height:32px; border-bottom:1px dotted #93bcc3; list-style-type:none; text-indent:8px; } .sub_menu ul li a { text-decoration:none; color:#FFFFFF; } .sub_menu ul li a.selected { background:url(http://www.cssblog.es/images/menu_selected.png) no-repeat; float:left; width:242px; height:32px; } .sub_menu ul li a:hover { background:url(http://www.cssblog.es/images/menu_selected.png) no-repeat; float:left; width:150px; height:30px; } - See more at: http://www.cssblog.es/disenando-un-bonito-menu-vertical-con-css/#sthash.AWv2bSbm.dpuf
- pagecontent
{
float: left; width: 620px; margin-left: 300px; min-height: 400px
} </style>
</html>
<html> <script type="text/x-mathjax-config"> MathJax.Hub.Config({tex2jax: {inlineMath: [['$','$'], ['\\(','\\)']]}}); MathJax.Hub.Config({ TeX: { equationNumbers: { autoNumber: "AMS" } } }); </script> <script type="text/javascript" src="http://cdn.mathjax.org/mathjax/latest/MathJax.js?config=TeX-AMS-MML_HTMLorMML"></script> <head> <title></title> </head> <body>
ENZYME
<img alt="" src="http://openwetware.org/images/e/e4/UANLEnzyme1.jpg" style="width: 410px; height: 385px;" />
Enzyme
In biological systems, chemical transformations are typically accelerated by enzymes, macromolecules capable of turning one or more compounds into others (substrates and products). The activity is determined greatly by their three-dimensional structure. Most enzymes are proteins, although several catalytic RNA molecules have been identified. They may also need to employ organic and inorganic cofactors for the reaction to occur. The process is based upon the diminishment of the activation energy needed for a reaction, greatly increasing its rate of reaction. The rate enhancement provided by these proteins can be as high as 10^19, while maintaining high substrate specificity.
Because of this, reaction rates are millions of times faster than un-catalyzed reactions. Enzymes are not consumed by the reactions that they take part in, and they do not alter the equilibrium. Enzyme activity can be affected by a wide variety of factors. Inhibitors and activators intervene directly in the reaction rate, environmental factors like temperature, pressure, pH and substrate concentration also play a part in these kinetics. For temperature and pH, usually exist a range of values for which the enzyme works better (optimal conditions). The enzyme activity lowers dramatically as you get farther away from this range of values. As for concentration, other kind of relationship is observed. With increasing concentration, enzyme activity increases, until we reach the most optimal performance. Further increase of concentration generally won’t have an impact on the enzyme activity.
Being able to determine these conditions allow us to manipulate the enzyme activity, thus achieving greater control over the reaction.
Michaelis-Menten kinetics
Michaelis-Menten kinetics is one of the oldest models for describing the catalytic activity of enzymes. The reaction cycle is divided into two basic steps: the reversible binding between the enzyme and substrate to form an intermediate complex, and the irreversible catalytic step to generate the product and release the enzyme; in which the first step is affected by the constants k1 and k-1, whereas the irreversible step only takes into account k2.
\begin{equation} E + S \leftrightarrow ES \rightarrow E^0 + P \end{equation}
The rate of consumption can be expressed by the formation of the ES complex in the following equation:
\begin{equation} \frac{d[ES]}{dt}=k_1[E][S]-k_{-1}[ES]-k_2[ES] \end{equation}
Using a steady-approximation and rearranging eq. 2 we obtain:
\begin{equation} [ES]= \frac{[E][S]}{K_M+[S]} \end{equation}
where ''KM'' is the Michaelis constant defined as
\begin{equation} K_M= \frac{k_{-1} + k_2}{k_1} \end{equation}
As it was mentioned in the introduction, single-enzyme studies have proven that the "traditional" enzyme kinetics do not apply, and a new approach is needed. Enzyme concentration is meaningless in a single-molecule level, so it is more appropriate to consider the probability ''PE(t)'' for the enzyme to find a catalytically active enzyme in a time ''t'' in the process. This is because the reaction is a stochastic event.
Therefore, the rate equations of each species are:
\begin{equation} \frac{d[E]}{dt}=-k_1[E][S]+k_{-1}[ES] \end{equation}
\begin{equation} \frac{d[ES]}{dt}=k_1[E][S]-(k_{-1}+k_2)[ES] \end{equation}
\begin{equation} \frac{d[E^0]}{dt}=\frac{d[P]}{dt}=k_2[ES] \end{equation} </body>
where ''t'' is the elapsed time, the initial conditions are [ES]=0 and [E<sup>0</sup>]=0 at ''t''=0. To derive the rate equations that describe the corresponding single-molecule Michaelis-Menten kinetics, the concentrations in equations 5-7 are replaced by the probabilities P of finding the single enzyme molecule in the states E, ES, and E<sup>0</sup> , leading to the equations:
\begin{equation} \frac{dP_E(t)}{dt}=-k_1^0P_E(t)+k_{-1}P_{ES}(t) \end{equation}
\begin{equation} \frac{dP_{ES}(t)}{dt}=k_1^0P_E(t)-(k_{-1}+k_2)P_{ES}(t) \end{equation}
\begin{equation} \frac{dP_E^0(t)}{dt}=k_2P_{ES}(t) \end{equation}
These equations must satisfy the conditions ''PE''(0)=1, ''PES''(0)=0 and ''PE0''=0 at ''t''=0 (start of the reaction). Also, ''PE''(t) + ''PES''(t) + ''PE0''(t)=1. The rate constant ''k''10 can be taken as ''k''10=''k''1[S], assuming [S] is time-independent. Given that a single enzyme is unlikely to deplete all the substrate presence, [S] can be considered constant, virtually being unaffected.
Equations 8-10 become a system of linear first-order differential equations that can be solved exactly for ''PE''(t), ''PES''(t) and ''PE0''(t).
Knowing ''PE0''(t), the probability that a turnover occurs between ''t'' and ''t +'' Δ''t'' is ''f(t)Δt, the same as Δ''PE0''(t). Taking this into account, in the limit of infinitesimal Δ''t'':
\begin{equation} f(t)=\frac{dP_E^0(t)}{dt}=k_2P_{ES}(t) \end{equation}
Solving equations 8-10, and using equation 11:
\begin{equation} f(t)=\frac{k_1k_2[S]}{2A}[exp(A+B)t-exp(B-A)t] \end{equation}
in which:
\begin{equation} A=\sqrt{(k_1[S]+k_{-1}+k_2)^2/4-k_1k_2[S]} \end{equation}
\begin{equation} B=- \frac{k_1[S]+k_{-1}+k_2}{2} \end{equation}
when the substrate concentration dependence [S] has been shown throught the relation ''k10 = k1''[S].
With these equations, we have used different values for ''k-1'' and [S], using values found in literature.
At the first moment of ''f(t)'', which would be the mean waiting time for the reaction, ''<t>'', and its reciprocal can be taken as the average reaction rate. Starting eq. 12:
\begin{equation} \frac{1}{\langle t \rangle)}=-\frac{(A^2-B^2)^2}{2Bk_1k_2[S]} \end{equation}
\begin{equation} \frac{1}{\langle t \rangle}=\frac{k_2[S]}{[S]+K_M} \end{equation}
Randomness parameter
The probability density ''f(t)'' completely characterizes single-enzyme kinetics, with the ''n''<sup>th</sup> moment being given by:
\begin{equation} f(t)= \int_0^\infty dt f(t)t^n \end{equation}
Although the first moment of ''f(t)'' can be described eq. 11, higher moments of ''f(t)'' are usually calculated along with a "randomness parameter". Implying no dynamic disorder, ''r'' is given by:
\begin{equation} r=\frac{(k_1[S]+k_2+k_{-1})^2-2k_1k_2[S]}{(k1[S]+k_2+k_{-1})^2} \end{equation}
As substrate concentration increases, r decreases, indicating the formation of an intermediate enzyme-substrate complex ES. At even higher concentrations, the catalytic step limits the reaction, as is the same when concentration is low and the substrate binding limits the rate.
Enzyme Reaction in a Container
Some reactions occur in spaces which have such a small volume that the normal approach to enzyme kinetics does not work. Even when taking into account probability instead of concentration, reaction cannot be modeled by aninfinite volume with a constant density. Examples of these systems include nanoparticles, zeolites or silica gels. In systems containing a small number of interacting species a relatively large fluctuationin the number of reactants is inherent. Therefore, the partitioning of the reagentspecies in small volume and the small number of the species present in eachindividual volume make it difficult to apply classical chemical kinetics to describe individual reactions in the container.
For a spherical system, in which molecules A and B react, the survival probability p(r,t) at the time t when A,B are at a d distance:
Enzyme Reaction in a Container
Some reactions occur in spaces which have such a small volume that the normal approach to enzyme kinetics does not work. Even when taking into account probability instead of concentration, reaction cannot be modeled by aninfinite volume with a constant density. Examples of these systems include nanoparticles, zeolites or silica gels. In systems containing a small number of interacting species a relatively large fluctuationin the number of reactants is inherent. Therefore, the partitioning of the reagentspecies in small volume and the small number of the species present in eachindividual volume make it difficult to apply classical chemical kinetics to describe individual reactions in the container.
For a spherical system, in which molecules A and B react, the survival probability p(r,t) at the time t when A,B are at a d distance:
\begin{equation}
\frac{δp(r,t)}{δt}=D\bigtriangledown^2p(r,t)
\end{equation}
\begin{equation} p(r,0)=1 \end{equation}
\begin{equation} p(d,t)=0 \end{equation}
\begin{equation} \bigg[ \frac{δp(r,t)}{δr} \bigg]_r=R =0 \end{equation}
These equations describe the random motion of A with respect to B, the system’s initial condition, distance needed to interact, and finally a boundary condition for the sphere.
In a general case where a sphere contains one B molecule and a number N of Amolecules that move mutually independently, the survival probability P(t) can bewritten as a product of the survival probabilities of all (A…B) pairs:
\begin{equation} P(t) = \prod_{i=1}^N p(r,t) \end{equation}
In this equation, riis the initial position of the ithA molecule. Averaging this last equation over therandom initial distribution of A molecules inside the sphere results in:
\begin{equation} P(r,t)= \bigg[\sum_{i=1}^\infty H_i•exp(-α_i^2 \frac{Dt}{R^2}\bigg]^N \end{equation}
\begin{equation} H_i=\frac{6\bigg(\frac{d}{R}\bigg)^2}{\alpha_i^2\bigg(1-\frac{d}{R}^2\bigg)\bigg[\frac{\alpha_i^2}{1+\alpha_i^2}-\frac{d}{R}^2\bigg]} \end{equation}
Thus, P(r,t) decays exponentially, indicating that the contained reaction obeys first-order kinetics.
RESULTS
Single-enzyme Michalis-Menten
After the solution of the differential equations system (eqs. 8-10), the analytical function of waiting time distribution f(t) was obtained. Then we introduced our parameters to understand the behavior of our specific system. The following constants obtained from literature:
\(k_1=10^7 M^{-1} s^{-1}\)
\(k_2=201 s^{-1}\)
\(k_{-1}=50-250 s^{-1}\)
\([S]=0.0001 - 0.1 mM\)
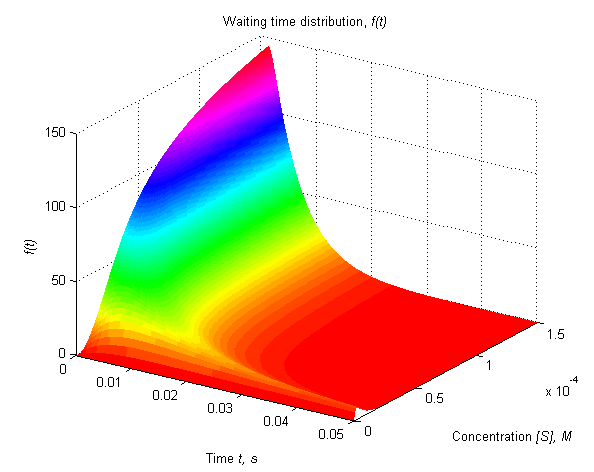
Using Matlab for calculating and drawing the plots for this data, we obtained the following figures, varying the values of [S]:
<img alt="" src="http://openwetware.org/images/2/2b/Waiting_time.png" style="width: 594px; height: 472px;" />
Fig.1 Waiting time distribution, as a function of time and [S]
At concentrations lower than 0.01 M, we can observe the lowest values of waiting time distributions. With increasing concentrations, the maximum contributions of waiting time distribution were observed around the first 0.02 seconds. After this time, their values drop dramatically, as we expected, because it is less probable to have a reaction after a longer elapsed time (because it must have already happened).
The same was done varying the values of k-1:
<img alt="" src="http://openwetware.org/images/c/c2/Waiting_time_2.png" />
Fig.2 Waiting time distribution, as a function of time and k-1
We can observe a change in the height of the curve´s peak aroung 0 and 0.02 seconds, as the values of k-1 increase. However, the same general behavior is observed for the waiting time distribution.
Then, with this function, we obtained the probability of survival (no reaction between the enzyme and the substrate) by the following equation:
\begin{equation}P(t)= 1- \int f(t) dt \end{equation}
<img alt="" src="http://openwetware.org/images/7/72/Waiting_prob_2D.png" style="width: 501px; height: 375px;" />
Fig.3 Probability of survival, as a function of time with varying [S] for the single-enzyme Michaelis-Menten model
As expected the probability drops dramatically as time increases, and drops faster for higher [S].
Confined space model
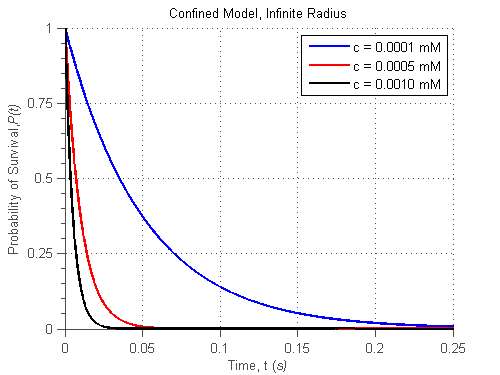
To prove the feasibility of the confined space model, we tested the special case of a container with an infinite radius, which has to behave as the previous model. For this, the probability of survival simplifies as follows:
\begin{equation} P(t)=exp(-4\pi cdDt)\end{equation}
where c is the concentration of the reactive species. During the reaction model, it was found that d = 1.6 x 1012 A2s-1. Again, using Matlab to compute these equations, we obtained the following figure:
<img alt="" src="http://openwetware.org/images/3/3c/Confined_inf.png" />
Fig.4 Probability of survival, as a function of time with varying [S] for the confined space model
The general behavior of this model is very similar to that of the single-enzyme Michaelis-Menten model, although the timescale of the former is in a different order of magnitude. We speculate that this is because the confined space model does is only taking into account geometric considerations (the reaction occurs when the molecules are close enough to each other). This model does not consider many other variables of the phenomenon, such as the location of active sites and substrate specificity, which are taken into account experimentaly by the rate of reaction coefficients in the single-enzyme Michaelis-Menten model. As such, it is apparent that the confined space model is a good model for this phenomenon, but needs to take into account more specific environmental conditions to fit with experimental data before applying it to confined spaces.
</html>

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}