Biomod/2013/Dresden/project
<html>
<link rel="stylesheet" href="http://fonts.googleapis.com/css?family=Lato:300,100&subset=latin"> <script src="//ajax.googleapis.com/ajax/libs/jquery/1.10.2/jquery.min.js" ></script>
<script type"text/javascript"> $(function () { $("style[media*='screen']").remove(); $("link[href*='favicon']").remove(); //fix heading var h1 = $(".firstHeading").text().split("/"); $(".firstHeading").text(h1[h1.length-1]); $("tr:odd").addClass("odd"); }); </script>
<style type="text/css"> /**** Base styles ****/
- column-one, #footer, div#sidebar-main, #contentSub, .firstHeading, #siteSub, #jump-to-nav, .printfooter{
display: none;
}
- content {
margin: 0; padding: 0;
background: #1A1A1A; border: medium none; }
html, body, div, span, object, iframe,
h1, h2, h3, h4, h5, h6, p, blockquote, pre,
abbr, address, cite, code, del, dfn, em, img, ins, kbd, q, samp,
small, strong, sub, sup, var, b, i, dl, dt, dd, ol, ul, li,
fieldset, form, label, legend,
table, caption, tbody, tfoot, thead, tr, th, td,
article, aside, canvas, details, figcaption, figure,
footer, header, hgroup, menu, nav, section, summary,
time, mark, audio, video {
margin: 0; padding: 0; border: 0; font: inherit; vertical-align: baseline;
}
article, aside, details, figcaption, figure, footer, header, hgroup, menu, nav, section {
display: block;
}
body {
/*padding: 15px;*/ font-family: 'Lato', 'Lucida Sans Regular', 'Lucida Grande', 'Lucida Sans Unicode', Arial, sans-serif; font-size: 2rem; font-weight: 200; line-height: 1.4; background: #1A1A1A; color: #999999 ; max-width: 1280px; padding-top: 55px; margin-left: auto; margin-right: auto;
} } h1, h2, h3, p, ul, ol, pre, dl {
margin-bottom: 1.0em; font-weight: 100;
}
h1, h2, #super-list, .box, .tagline, #index-list {
font-family: 'Lato', 'Helvetica Neue', Arial, sans-serif;
}
h1, h2, h3 { font-weight: 300; }
h1 {
font-size: 32px; line-height: 1.1em;
}
h2 {
font-size: 48px; line-height: 1.1em;
}
a, a code {
color: #FB4; text-decoration: none;
}
a:hover, a:hover code {
color: #4BF;
}
a:active, a:active code {
color: #1FB; /*background: black;*/
}
a img { border: none; }
a.anchor{display: block; position: relative; top: -75px; visibility: hidden;}
p{ text-align:left; }
em { color: #00EF00; } strong { font-weight: bold; }
blockquote {
padding-left: 1.0em; margin-left: 1.0em; border-left: 1px solid #333; font-style: italic;
}
- scrollUp {
background: url("http://openwetware.org/images/8/87/Top.png") no-repeat scroll 0 0 transparent;
bottom: 20px;
height: 38px;
right: 10px;
width: 38px;
{kind=link}
}
nav {
background: rgba(25, 25, 25, 0.85); padding: 0px; position: fixed; top: 0px; left: 0px; right: 0px; z-index: 100;
}
nav ul {
width: 100%; margin: 0px auto; padding: 0px; list-style-type: none;
} nav ul li {
float: left; line-height:2.5;
} nav ul li.selected {
border-bottom: 10px solid #d30d15;
}
nav ul li.home {
padding: 10px;
} nav ul a {
float: left; text-decoration: none; color: #F2F2F2; text-transform: uppercase; font-size: 20px; font-weight: 300; padding: 10px 20px 0;
}
- subnav {
background-color: rgba(25, 25, 25, 0.8);
border-radius: 0 20px 20px 0; left:0px;
font-size: 18px; font-weight: 300; line-height: 1.8; list-style: none outside none; margin-left: 0; padding: 20px;
z-index: 10; }
- subnav li {
list-style: none outside none;
}
- content {
margin-top: 60px;
}
- filters > li{
margin: 5px; display: inline-block; }
.box.clickable:hover{ background: none repeat scroll 0 0 #fff; }
.clickable img { transition: 0.3s ease; }
.clickable img:hover { opacity:0.9; transition: 0.3s ease; }
.background { left: 0;
margin: 0; max-width: 100%; padding: 0; position: absolute;
} /*the boxes*/
.box.b2x2{ height: 460px; width: 460px; } .box.b2x2 > img{ display: block;
margin-left: auto; margin-right: auto; margin-top: 10px; width: 400px;
}
.box.b2x1{ height: 460px; }
.box.b1x2{ width: 460px; }
.box.b1x3{ width: 700px; }
/*start page*/
.box.intro { font-size:7.2rem;}
.box > p { font-size: 16px;
padding: 0 20px; margin-top: 10px; text-align: justify;
font-weight: 300; }
.box > h2 { font-size: 40px;
font-weight: 300;
text-align:left; margin-left: 20px;
margin-top: 15px;
}
.tease > h2 {
font-size: 80px; font-weight: 100; margin-top: 80px;
}
/*project*/
.project{ background-attachment: fixed;
width: 100%;
padding-bottom: 40px; }
.project h2 { color: #FFFFFF;
font-weight: 300; margin-bottom: 30px; margin-left: 180px; padding-top: 20px; position: relative;
}
.project h3 {
font-size: 26px;
}
.project .box { margin-bottom: 20px;
margin-top: 0;
}
.interlude{ background: none repeat scroll 0 0 #2A2A2A; box-shadow: 0 0 25px rgba(0, 0, 0, 0.8); border: 1px solid rgba(0, 0, 0, 0.3);
height: 150px; position: relative; z-index: 3;
}
.interlude *{ float:left; }
.interlude h2{ color: #FFFFFF;
display: block; font-weight: 300; line-height: 150px; margin-left: 24%; margin-right: -14%; width: 50%;
}
.interlude img{ float: left;
line-height: 150px; margin-left: 10%; margin-top: 25px; vertical-align: middle;
}
.clear{
clear: both;
}
- motivation{ background-image: url("http://openwetware.org/images/d/d9/Motivation.png");}
{kind=link}
- vesicle{ background-image: url("http://openwetware.org/images/0/0e/Vesicle_large_final.png");}
{kind=link}
- origami{ background-image: url("http://openwetware.org/images/8/89/DNA_background_final.png");}
{kind=link}
- reaction{ background-image: url("http://openwetware.org/images/c/c3/IR_background.png");}
{kind=link}
- nanoreactor{ background-image: url("http://openwetware.org/images/a/a5/Nanocontainer_background.png");}
{kind=link}
- applications{ background-image: url("http://openwetware.org/images/f/fa/Application_background.png");}
{kind=link}
- future{ background-image: url("http://openwetware.org/images/8/83/Future_appl_background.png");}
{kind=link}
.project_box{ background: none repeat scroll 0 0 #FFFFFF;
color: #000000; display: block; float: left; font-size: 18px; font-weight: 300; line-height: 1.6; margin-left: 180px; margin-right: auto; overflow: hidden; padding: 10px; width: 50%;
}
.figure_box{
display: block; float: left; margin-left: 20px; overflow: hidden; width: 230px;
}
/*se*/
.project_box h2{ color: #1A1A1A;
display: block; font-weight: 300;
margin-left: 0%; margin-right: -10%; width: 90%;
}
.project_box p{
text-align: justify;
margin-bottom: 18px; }
.project_box li { margin-left:50px }
- pb_mot.project_box{
height: 350px; }
- pb_dna_scaff.project_box{
height: 400px; }
- pb_dna_req.project_box{
height: 250px; }
- pb_poly_intro.project_box{
/*right: -20%;*/ height: 200px;
}
- pb_poly_pmoxa.project_box{
/*right: -20%;*/ height: 600px; width: 1000px; overflow:scroll; }
- pb_ir.project_box{
/*right: -20%;*/ height: 1300px; }
- pb_poly_cp.project_box{
/*right: -20%;*/ height: 600px; }
/*team page*/ .bio_box {
background: none repeat scroll 0 0 #E74C3C; float: left; font-size: 15px; height: 440px; padding: 15px; text-align: justify; width: 210px;
}
.bio_box > .name{ font-size: 24px;
font-weight: 300; margin-bottom: 25px; text-align: center; width: 100%;
}
.bio_box > p{
text-align: justify;
font-weight: 300; font-size: 16px; } .box.big img{
opacity:1;
}
.flag > *{ float:left; } .flag > p{ font-size: 18px;
position: relative; text-align: center; top: -6px; width: 75%;
margin-bottom: 10px; }
- team .big{
opacity:1; }
.head{ width:220px; float:left; }
/*sponsor page*/
- sponsors .box {
background: none repeat scroll 0 0 white;
}
- sponsors figcaption {
height: 65px; width: 100%;
font-size: 15px;
font-weight: 300; top: auto; bottom: 0; opacity: 0; transform: translateY(100%);
transition: transform 0.4s, opacity 0.1s 0.3s; -webkit-transform: translateY(100%);
-webkit-transition: -webkit-transform 0.4s, opacity 0.1s 0.3s;
}
- sponsors .descr{
background: none repeat scroll 0 0 rgba(0, 0, 0, 0.4);
font-size: 12px; font-weight: 300; height: 60px;
margin: 0;
padding-left: 10px; padding-right: 10px; padding-top: 10px; text-align: justify; top: -155px; line-height: 1.3;
}
- sponsors .descr p{
width:90%; margin-left:auto; margin-right:auto; }
- sponsors figure.clickable:hover figcaption{
opacity: 1; transform: translateY(0px); transition: transform 0.4s, opacity 0.1s;
-webkit-transform: translateY(0px);
-webkit-transition: -webkit-transform 0.4s, opacity 0.1s;
}
- sponsors figure:hover .descr{
opacity: 1; transform: translateY(155px); transition: transform 0.4s, opacity 0.1s;
-webkit-transform: translateY(155px); -webkit-transition: -webkit-transform 0.4s, opacity 0.1s; }
/*gallery*/
- gallery .box img{
min-height: 220px;
min-width: 220px;
} /*ptocols*/ .protocol_box{ background: none repeat scroll 0 0 #FFFFFF;
color: #000000; display: block; float: left; font-size: 18px; font-weight: 300; line-height: 1.6; margin-left: 40px; margin-right: auto; overflow: hidden; padding: 10px; width: 66%;
} .protocol_box li { margin-left:50px }
.protocol_box p{
text-align: justify;
margin-top: 18px; }
.protocol_box h1 { font-size: 30px; }
.protocol_box h2 { font-size: 24px; }
.protocol_box h3 { font-size: 22px; }
/*Outreach*/
.outreach_box{ background: none repeat scroll 0 0 #FFFFFF;
color: #000000; display: block; float: left; font-size: 18px; font-weight: 300; line-height: 1.6; margin-left: 10px; margin-right: auto; overflow: hidden; padding: 10px; width: 70%;
}
.outreach_box li { margin-left:50px }
/*Acknowlegement*/
.ack_box{ background: none repeat scroll 0 0 #FFFFFF;
color: #000000;
text-align: center;
display: block; float: left; font-size: 18px; font-weight: 300; line-height: 1.6; margin-left: 10px; margin-right: auto; overflow: hidden; padding: 10px; width: 80%;
} .ack_box p { text-align: center; } .next, .prev{ z-index: 99; background-image: url("http://openwetware.org/images/5/55/Fancybox_sprite.png"); width: 36px; height: 36px; top: 200px; }
{kind=link}
figure.box > .next {
left: 425px;
background-position: 0 -72px;
}
figure.box > .prev { background-position: 0 -36px; }
/**** Isotope styles ****/
/* required for containers to inherit vertical size from window */ html, body {
height: 100%;
}
- container {
padding: 5px; margin-bottom: 20px;
}
.box {
width: 220px; height: 220px; margin: 10px; float: left; overflow: hidden; position: relative; background: #2A2A2A; color: #F2F2F2; display: table-cell; text-align: center; vertical-align: middle; overflow:hidden;
}
figure.box > *{ left: 0;
position: absolute; right: 0;
}
.box figure{ overflow: hidden; }
.box figcaption {
background: none repeat scroll 0 0 rgba(0, 0, 0, 0.4); bottom: 0; font-size: 20px;
font-weight: 300;
padding-left: 5px; text-align: center; width: 100%;
z-index: 4; }
.clickable .box:hover {
cursor: pointer;
}
/* The Magnificent Clearfix: nicolasgallagher.com/micro-clearfix-hack/ */ .clearfix:before, .clearfix:after { content: ""; display: table; } .clearfix:after { clear: both; } .clearfix { zoom: 1; }
/* Start: Recommended Isotope styles */
/**** Isotope Filtering ****/
.isotope-item {
z-index: 2;
}
.isotope-hidden.isotope-item {
pointer-events: none; z-index: 1;
}
/**** Isotope CSS3 transitions ****/
.isotope, .isotope .isotope-item {
-webkit-transition-duration: 0.8s;
-moz-transition-duration: 0.8s;
-ms-transition-duration: 0.8s;
-o-transition-duration: 0.8s;
transition-duration: 0.8s;
}
.isotope {
-webkit-transition-property: height, width;
-moz-transition-property: height, width;
-ms-transition-property: height, width;
-o-transition-property: height, width;
transition-property: height, width;
}
.isotope .isotope-item {
-webkit-transition-property: -webkit-transform, opacity;
-moz-transition-property: -moz-transform, opacity;
-ms-transition-property: -ms-transform, opacity;
-o-transition-property: -o-transform, opacity;
transition-property: transform, opacity;
} .rs-wrap:after, .rs-slider:after, .rs-thumb-wrap:after, .rs-arrows:after, .rs-caption:after {
content: "."; display: block; height: 0; clear: both; line-height: 0; visibility: hidden;
}
/* ===[ Slider ]=== */
.rs-wrap {
position: relative; max-width: 100%;
}
.rs-slide-bg { *zoom: 1 }
.rs-slider > li > a { display: block }
.rs-slider > li {
list-style: none; filter: alpha(opacity=0); opacity: 0; width: 100%; height: 100%; margin: 0 -100% 0 0; padding: 0; float: left; position: relative;
}
.rs-slider > li > a {
padding: 0;
background: none;
-webkit-border-radius: 0;
-moz-border-radius: 0;
border-radius: 0;
}
.rs-slider > li img {
display: block;
max-width: 100%;
max-height: 100%;
-ms-interpolation-mode: bicubic;
}
/* ===[ Thumbnails ]=== */
.rs-thumb-wrap { *zoom: 1 }
.rs-thumb-wrap > a {
display: block;
float: left;
position: relative;
-moz-box-sizing: border-box;
-webkit-box-sizing: border-box;
box-sizing: border-box;
-webkit-backface-visibility: hidden; /* Hardware accelerate to prevent jumps on transition */
}
.rs-thumb-wrap > a > img {
max-width: 100%;
max-height: 100%;
display: block;
-ms-interpolation-mode: bicubic;
}
.rs-thumb-wrap > a:first-child { margin-left: 0!important }
/* ===[ Arrows ]=== */
.rs-arrows .rs-next, .rs-arrows .rs-prev { z-index: 1; background-image: url("fancybox_sprite.png");}
.rs-arrows .rs-next, .rs-arrows .rs-prev { z-index: 1; background-image: url("fancybox_sprite.png");}
.rs-arrows:hover .rs-next, .rs-arrows:hover .rs-prev { z-index: 2; }
/* ===[ Captions ]=== */
.rs-caption {
position: absolute; max-height: 100%; overflow: auto; -moz-box-sizing: border-box; -webkit-box-sizing: border-box; box-sizing: border-box; bottom: 0; left: 0;
}
.rs-caption.rs-top-left {
top: 0; bottom: auto;
}
.rs-caption.rs-top-right {
top: 0; right: 0; left: auto; bottom: auto;
}
.rs-caption.rs-bottom-left {
bottom: 0; left: 0;
}
.rs-caption.rs-bottom-right {
right: 0; left: auto; border-bottom: none; border-right: none;
}
.rs-caption.rs-top {
top: 0; bottom: auto; width: 100%!important;
}
.rs-caption.rs-bottom { width: 100%!important }
.rs-caption.rs-left {
top: 0; height: 100%;
}
.rs-caption.rs-right {
top: 0; left: auto; right: 0; height: 100%;
}
/* ===[ Grid ]=== */
.rs-grid {
position: absolute; overflow: hidden; width: 100%; height: 100%; display: none;
}
.rs-gridlet {
position: absolute; opacity: 1;
}
/* Optional - remove captions at smaller screen widths @media screen and (max-width: 480px) { .rs-caption { opacity: 0!important; } }
- /
.project_box > img {
margin-left: 90px;
}
- protocols, #polymers_protocols, #origami_protocols, #reaction_protocols, #nanocontainer_protocols, #imaging_protocols {
font-size: 20px; font-weight: 300; margin-bottom: 30px; margin-left: 50px;
}
- protocols > h2, #polymers_protocols > h2, #origami_protocols > h2, #reaction_protocols > h2, #nanocontainer_protocols > h2, #imaging_protocols > h2 {
margin-bottom: 20px; margin-top: 20px;
}
- protocols .interlude, #polymers_protocols .interlude, #origami_protocols .interlude, #reaction_protocols .interlude, #nanocontainer_protocols .interlude, #imaging_protocols .interlude {
margin-left: -50px !important;
}
- protocols > ul {
margin-bottom: 30px; margin-left: 30px; margin-top: 20px;
}
li > ul {
margin-left: 20px;
}
/*achievement*/
.achievement_box{ background: none repeat scroll 0 0 #FFFFFF;
color: #000000; display: block; float: left; font-size: 18px; font-weight: 300; line-height: 1.6; margin-left: 180px; margin-right: auto; overflow: hidden; padding: 10px; width: 50%;
} @-webkit-keyframes glowing {
/* line 8, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 9, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 10, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
@-khtml-keyframes glowing {
/* line 14, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 15, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 16, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
@-moz-keyframes glowing {
/* line 20, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 21, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 22, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
@-ms-keyframes glowing {
/* line 26, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 27, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 28, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
@-o-keyframes glowing {
/* line 32, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 33, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 34, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
@keyframes glowing {
/* line 38, ../scss/partials/_glow.scss */
from {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 39, ../scss/partials/_glow.scss */
50% {
-webkit-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 16px rgba(44, 154, 219, 0.8), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
/* line 40, ../scss/partials/_glow.scss */
to {
-webkit-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
-moz-box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
box-shadow: 0px 0px 0px rgba(44, 154, 219, 0.3), 0px 1px 2px rgba(0, 0, 0, 0.2);
}
}
/* line 25, ../scss/partials/_buttons.scss */ .button {
-webkit-box-shadow: inset 0px 1px 0px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.15); -moz-box-shadow: inset 0px 1px 0px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.15); box-shadow: inset 0px 1px 0px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.15); background-color: #eeeeee; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #fbfbfb), color-stop(100%, #e1e1e1)); background: -webkit-linear-gradient(top, #fbfbfb, #e1e1e1); background: -moz-linear-gradient(top, #fbfbfb, #e1e1e1); background: -o-linear-gradient(top, #fbfbfb, #e1e1e1); background: linear-gradient(top, #fbfbfb, #e1e1e1); display: -moz-inline-stack; display: inline-block; vertical-align: middle; *vertical-align: auto; zoom: 1; *display: inline; border: 1px solid #d4d4d4; height: 32px; line-height: 32px; padding: 0px 25.6px; font-weight: 300; font-size: 14px; font-family: "HelveticaNeue-Light", "Helvetica Neue Light", "Helvetica Neue", Helvetica, Arial, "Lucida Grande", sans-serif; color: #666666; text-shadow: 0 1px 1px white; margin: 0; text-decoration: none; text-align: center;
} /* line 43, ../scss/partials/_buttons.scss */ .button:hover {
background-color: #eeeeee; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #ffffff), color-stop(100%, #dcdcdc)); background: -webkit-linear-gradient(top, #ffffff, #dcdcdc); background: -moz-linear-gradient(top, #ffffff, #dcdcdc); background: -o-linear-gradient(top, #ffffff, #dcdcdc); background: linear-gradient(top, #ffffff, #dcdcdc);
} /* line 47, ../scss/partials/_buttons.scss */ .button:active {
-webkit-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white; -moz-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white; box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white; text-shadow: 0px 1px 0px rgba(255, 255, 255, 0.4); background: #eeeeee; color: #bbbbbb;
} /* line 53, ../scss/partials/_buttons.scss */ .button:focus {
outline: none;
}
/* line 59, ../scss/partials/_buttons.scss */ input.button, button.button {
height: 34px; cursor: pointer;
}
/* line 65, ../scss/partials/_buttons.scss */ .button-block {
display: block;
}
/* line 74, ../scss/partials/_buttons.scss */ .button.disabled, .button.disabled:hover, .button.disabled:active, input.button:disabled, button.button:disabled {
-webkit-box-shadow: 0px 1px 2px rgba(0, 0, 0, 0.1); -moz-box-shadow: 0px 1px 2px rgba(0, 0, 0, 0.1); box-shadow: 0px 1px 2px rgba(0, 0, 0, 0.1); background: #EEE; border: 1px solid #DDD; text-shadow: 0 1px 1px white; color: #CCC; cursor: default;
}
/* line 84, ../scss/partials/_buttons.scss */ .button-wrap {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #e3e3e3), color-stop(100%, #f2f2f2)); background: -webkit-linear-gradient(top, #e3e3e3, #f2f2f2); background: -moz-linear-gradient(top, #e3e3e3, #f2f2f2); background: -o-linear-gradient(top, #e3e3e3, #f2f2f2); background: linear-gradient(top, #e3e3e3, #f2f2f2); -webkit-border-radius: 200px; -moz-border-radius: 200px; -ms-border-radius: 200px; -o-border-radius: 200px; border-radius: 200px; -webkit-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.04); -moz-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.04); box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.04); padding: 10px; display: inline-block;
}
/* line 100, ../scss/partials/_buttons.scss */ .button-rounded {
-webkit-border-radius: 3px; -moz-border-radius: 3px; -ms-border-radius: 3px; -o-border-radius: 3px; border-radius: 3px;
}
/* line 100, ../scss/partials/_buttons.scss */ .button-pill {
-webkit-border-radius: 50px; -moz-border-radius: 50px; -ms-border-radius: 50px; -o-border-radius: 50px; border-radius: 50px;
}
/* line 100, ../scss/partials/_buttons.scss */ .button-circle {
-webkit-border-radius: 240px; -moz-border-radius: 240px; -ms-border-radius: 240px; -o-border-radius: 240px; border-radius: 240px; -webkit-box-shadow: inset 0px 1px 1px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.2); -moz-box-shadow: inset 0px 1px 1px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.2); box-shadow: inset 0px 1px 1px rgba(255, 255, 255, 0.5), 0px 1px 2px rgba(0, 0, 0, 0.2); width: 120px; line-height: 120px; height: 120px; padding: 0px; border-width: 4px; font-size: 18px;
}
/* line 135, ../scss/partials/_buttons.scss */ .button-primary {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #00b5e5), color-stop(100%, #008db2)); background: -webkit-linear-gradient(top, #00b5e5, #008db2); background: -moz-linear-gradient(top, #00b5e5, #008db2); background: -o-linear-gradient(top, #00b5e5, #008db2); background: linear-gradient(top, #00b5e5, #008db2); background-color: #00a1cb; border-color: #007998; color: white; text-shadow: 0 -1px 1px rgba(0, 40, 50, 0.35);
} /* line 142, ../scss/partials/_buttons.scss */ .button-primary:hover {
background-color: #00a1cb; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #00c9fe), color-stop(100%, #008db2)); background: -webkit-linear-gradient(top, #00c9fe, #008db2); background: -moz-linear-gradient(top, #00c9fe, #008db2); background: -o-linear-gradient(top, #00c9fe, #008db2); background: linear-gradient(top, #00c9fe, #008db2);
} /* line 146, ../scss/partials/_buttons.scss */ .button-primary:active {
background: #1495b7; color: #005065;
}
/* line 135, ../scss/partials/_buttons.scss */ .button-action {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #8fcf00), color-stop(100%, #6b9c00)); background: -webkit-linear-gradient(top, #8fcf00, #6b9c00); background: -moz-linear-gradient(top, #8fcf00, #6b9c00); background: -o-linear-gradient(top, #8fcf00, #6b9c00); background: linear-gradient(top, #8fcf00, #6b9c00); background-color: #7db500; border-color: #5a8200; color: white; text-shadow: 0 -1px 1px rgba(19, 28, 0, 0.35);
} /* line 142, ../scss/partials/_buttons.scss */ .button-action:hover {
background-color: #7db500; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #a0e800), color-stop(100%, #6b9c00)); background: -webkit-linear-gradient(top, #a0e800, #6b9c00); background: -moz-linear-gradient(top, #a0e800, #6b9c00); background: -o-linear-gradient(top, #a0e800, #6b9c00); background: linear-gradient(top, #a0e800, #6b9c00);
} /* line 146, ../scss/partials/_buttons.scss */ .button-action:active {
background: #76a312; color: #374f00;
}
/* line 135, ../scss/partials/_buttons.scss */ .button-highlight {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #fa9915), color-stop(100%, #d87e04)); background: -webkit-linear-gradient(top, #fa9915, #d87e04); background: -moz-linear-gradient(top, #fa9915, #d87e04); background: -o-linear-gradient(top, #fa9915, #d87e04); background: linear-gradient(top, #fa9915, #d87e04); background-color: #f18d05; border-color: #bf7004; color: white; text-shadow: 0 -1px 1px rgba(91, 53, 2, 0.35);
} /* line 142, ../scss/partials/_buttons.scss */ .button-highlight:hover {
background-color: #f18d05; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #fba42e), color-stop(100%, #d87e04)); background: -webkit-linear-gradient(top, #fba42e, #d87e04); background: -moz-linear-gradient(top, #fba42e, #d87e04); background: -o-linear-gradient(top, #fba42e, #d87e04); background: linear-gradient(top, #fba42e, #d87e04);
} /* line 146, ../scss/partials/_buttons.scss */ .button-highlight:active {
background: #d8891e; color: #8d5303;
}
/* line 135, ../scss/partials/_buttons.scss */ .button-caution {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #e8543f), color-stop(100%, #d9331a)); background: -webkit-linear-gradient(top, #e8543f, #d9331a); background: -moz-linear-gradient(top, #e8543f, #d9331a); background: -o-linear-gradient(top, #e8543f, #d9331a); background: linear-gradient(top, #e8543f, #d9331a); background-color: #e54028; border-color: #c22d18; color: white; text-shadow: 0 -1px 1px rgba(103, 24, 13, 0.35);
} /* line 142, ../scss/partials/_buttons.scss */ .button-caution:hover {
background-color: #e54028; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #eb6855), color-stop(100%, #d9331a)); background: -webkit-linear-gradient(top, #eb6855, #d9331a); background: -moz-linear-gradient(top, #eb6855, #d9331a); background: -o-linear-gradient(top, #eb6855, #d9331a); background: linear-gradient(top, #eb6855, #d9331a);
} /* line 146, ../scss/partials/_buttons.scss */ .button-caution:active {
background: #cd5240; color: #952312;
}
/* line 135, ../scss/partials/_buttons.scss */ .button-royal {
background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #99389f), color-stop(100%, #752a79)); background: -webkit-linear-gradient(top, #99389f, #752a79); background: -moz-linear-gradient(top, #99389f, #752a79); background: -o-linear-gradient(top, #99389f, #752a79); background: linear-gradient(top, #99389f, #752a79); background-color: #87318c; border-color: #632466; color: white; text-shadow: 0 -1px 1px rgba(26, 9, 27, 0.35);
} /* line 142, ../scss/partials/_buttons.scss */ .button-royal:hover {
background-color: #87318c; background: -webkit-gradient(linear, 50% 0%, 50% 100%, color-stop(0%, #ab3eb2), color-stop(100%, #752a79)); background: -webkit-linear-gradient(top, #ab3eb2, #752a79); background: -moz-linear-gradient(top, #ab3eb2, #752a79); background: -o-linear-gradient(top, #ab3eb2, #752a79); background: linear-gradient(top, #ab3eb2, #752a79);
} /* line 146, ../scss/partials/_buttons.scss */ .button-royal:active {
background: #764479; color: #3e1740;
}
/* line 160, ../scss/partials/_buttons.scss */ .button-flat {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #eeeeee; border: none; text-shadow: none;
} /* line 169, ../scss/partials/_buttons.scss */ .button-flat:hover {
background: #fbfbfb;
} /* line 172, ../scss/partials/_buttons.scss */ .button-flat:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #eeeeee; color: #bbbbbb;
} /* line 177, ../scss/partials/_buttons.scss */ .button-flat.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 189, ../scss/partials/_buttons.scss */ .button-flat-primary {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #00a1cb; color: white; text-shadow: none; border: none;
} /* line 198, ../scss/partials/_buttons.scss */ .button-flat-primary:hover {
background: #00b5e5;
} /* line 201, ../scss/partials/_buttons.scss */ .button-flat-primary:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #1495b7; color: #00647f;
} /* line 206, ../scss/partials/_buttons.scss */ .button-flat-primary.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 189, ../scss/partials/_buttons.scss */ .button-flat-action {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #7db500; color: white; text-shadow: none; border: none;
} /* line 198, ../scss/partials/_buttons.scss */ .button-flat-action:hover {
background: #8fcf00;
} /* line 201, ../scss/partials/_buttons.scss */ .button-flat-action:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #76a312; color: #486900;
} /* line 206, ../scss/partials/_buttons.scss */ .button-flat-action.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 189, ../scss/partials/_buttons.scss */ .button-flat-highlight {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #f18d05; color: white; text-shadow: none; border: none;
} /* line 198, ../scss/partials/_buttons.scss */ .button-flat-highlight:hover {
background: #fa9915;
} /* line 201, ../scss/partials/_buttons.scss */ .button-flat-highlight:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #d8891e; color: #a66103;
} /* line 206, ../scss/partials/_buttons.scss */ .button-flat-highlight.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 189, ../scss/partials/_buttons.scss */ .button-flat-caution {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #e54028; color: white; text-shadow: none; border: none;
} /* line 198, ../scss/partials/_buttons.scss */ .button-flat-caution:hover {
background: #e8543f;
} /* line 201, ../scss/partials/_buttons.scss */ .button-flat-caution:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #cd5240; color: #ac2815;
} /* line 206, ../scss/partials/_buttons.scss */ .button-flat-caution.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 189, ../scss/partials/_buttons.scss */ .button-flat-royal {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none; -webkit-transition-property: background; -moz-transition-property: background; -o-transition-property: background; transition-property: background; -webkit-transition-duration: 0.3s; -moz-transition-duration: 0.3s; -o-transition-duration: 0.3s; transition-duration: 0.3s; background: #87318c; color: white; text-shadow: none; border: none;
} /* line 198, ../scss/partials/_buttons.scss */ .button-flat-royal:hover {
background: #99389f;
} /* line 201, ../scss/partials/_buttons.scss */ .button-flat-royal:active {
-webkit-transition-duration: 0s; -moz-transition-duration: 0s; -o-transition-duration: 0s; transition-duration: 0s; background: #764479; color: #501d53;
} /* line 206, ../scss/partials/_buttons.scss */ .button-flat-royal.disabled {
-webkit-box-shadow: none; -moz-box-shadow: none; box-shadow: none;
}
/* line 221, ../scss/partials/_buttons.scss */ .button-large {
font-size: 19px; height: 38.4px; line-height: 38.4px; padding: 0px 30.72px;
}
/* line 245, ../scss/partials/_buttons.scss */ input.button-large, button.button-large {
height: 40.4px;
}
/* line 221, ../scss/partials/_buttons.scss */ .button-small {
font-size: 16px; height: 25.6px; line-height: 25.6px; padding: 0px 20.48px;
}
/* line 245, ../scss/partials/_buttons.scss */ input.button-small, button.button-small {
height: 27.6px;
}
/* line 221, ../scss/partials/_buttons.scss */ .button-tiny {
font-size: 12px; height: 22.4px; line-height: 22.4px; padding: 0px 17.92px;
}
/* line 245, ../scss/partials/_buttons.scss */ input.button-tiny, button.button-tiny {
height: 24.4px;
}
/* line 265, ../scss/partials/_buttons.scss */ .button.glow {
-webkit-animation-duration: 3s; -moz-animation-duration: 3s; -ms-animation-duration: 3s; -o-animation-duration: 3s; animation-duration: 3s; -webkit-animation-iteration-count: infinite; -khtml-animation-iteration-count: infinite; -moz-animation-iteration-count: infinite; -ms-animation-iteration-count: infinite; -o-animation-iteration-count: infinite; animation-iteration-count: infinite; -webkit-animation-name: glowing; -khtml-animation-name: glowing; -moz-animation-name: glowing; -ms-animation-name: glowing; -o-animation-name: glowing; animation-name: glowing;
} /* line 268, ../scss/partials/_buttons.scss */ .button.glow:active {
-webkit-animation-name: none; -moz-animation-name: none; -ms-animation-name: none; -o-animation-name: none; animation-name: none; -webkit-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white; -moz-box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white; box-shadow: inset 0px 1px 3px rgba(0, 0, 0, 0.3), 0px 1px 0px white;
}
/* line 279, ../scss/partials/_buttons.scss */ .button-dropdown {
position: relative; overflow: visible; display: inline-block;
} /* line 284, ../scss/partials/_buttons.scss */ .button-dropdown .button-overlay {
position: fixed; top: 0px; left: 0px; right: 0px; bottom: 0px; z-index: 999;
} /* line 295, ../scss/partials/_buttons.scss */ .button-dropdown .button .icon-caret-down {
font-size: 90%; margin: 0px 0px 0px 3px; vertical-align: middle;
} /* line 302, ../scss/partials/_buttons.scss */ .button-dropdown ul.button-dropdown-menu-below {
top: 115%;
} /* line 305, ../scss/partials/_buttons.scss */ .button-dropdown ul.button-dropdown-menu-above {
bottom: 115%; top: auto;
} /* line 311, ../scss/partials/_buttons.scss */ .button-dropdown ul {
-webkit-box-shadow: 0px 3px 6px rgba(0, 0, 0, 0.6); -moz-box-shadow: 0px 3px 6px rgba(0, 0, 0, 0.6); box-shadow: 0px 3px 6px rgba(0, 0, 0, 0.6); -webkit-border-radius: 3px; -moz-border-radius: 3px; -ms-border-radius: 3px; -o-border-radius: 3px; border-radius: 3px; display: none; position: absolute; background: #fcfcfc; top: -2px; left: -2px; z-index: 1000; padding: 0px; margin: 0px; list-style-type: none; min-width: 102%;
} /* line 325, ../scss/partials/_buttons.scss */ .button-dropdown ul li {
padding: 0px; margin: 0px; display: block;
} /* line 330, ../scss/partials/_buttons.scss */ .button-dropdown ul li:first-child a {
-moz-border-radius-topleft: 3px; -webkit-border-top-left-radius: 3px; border-top-left-radius: 3px; -moz-border-radius-topright: 3px; -webkit-border-top-right-radius: 3px; border-top-right-radius: 3px;
} /* line 333, ../scss/partials/_buttons.scss */ .button-dropdown ul li:last-child a {
-moz-border-radius-bottomleft: 3px; -webkit-border-bottom-left-radius: 3px; border-bottom-left-radius: 3px; -moz-border-radius-bottomright: 3px; -webkit-border-bottom-right-radius: 3px; border-bottom-right-radius: 3px;
} /* line 339, ../scss/partials/_buttons.scss */ .button-dropdown ul .button-dropdown-divider {
-webkit-box-shadow: inset 0px 1px 0px white; -moz-box-shadow: inset 0px 1px 0px white; box-shadow: inset 0px 1px 0px white; border-top: 1px solid #e4e4e4;
} /* line 344, ../scss/partials/_buttons.scss */ .button-dropdown ul a {
display: block; padding: 0px 20px; text-decoration: none; font-size: 12px; color: #333333; line-height: 30px; white-space: nowrap;
} /* line 353, ../scss/partials/_buttons.scss */ .button-dropdown ul a:hover {
background-color: #3c6ab9; color: white;
}
- subnav-sticky-wrapper {
height: 5px !important;
}
table {
border-collapse: collapse; margin: auto auto 40px; width: 635px;;
} th {
background-color: #5F5F5F; border: 1px solid #999999; color: #FFFFFF;
} tr td {
border: 1px solid #999999; text-align: center;
} tr.odd td {
background-color: #EEEEEE; color: #000000;
} .ref li {
font-size: 14px; font-weight: 300;
} </style> <link href="http://openwetware.org/images/2/29/Nano_icon.png" rel="shortcut icon">
{kind=link}
<nav role="navigation">
{kind=link}
</nav> <script src="https://biomod2013.googlecode.com/svn/trunk/js/jquery.isotope.min.js"></script> <script src="https://biomod2013.googlecode.com/svn/trunk/js/jquery.refineslide.min.js"></script>
<script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/fb/jquery.fancybox.pack.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/fb/helpers/jquery.fancybox-buttons.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/fb/helpers/jquery.fancybox-media.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/fb/helpers/jquery.fancybox-thumbs.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.easing.min.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.scrollUp.min.js"></script>
<script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.stellar.min.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.sticky.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.scrollTo.min.js"></script> <script type="text/javascript" src="http://biomod2013.googlecode.com/svn/trunk/js/jquery.localscroll.min.js"></script>
<script>
(function(i,s,o,g,r,a,m){i['GoogleAnalyticsObject']=r;i[r]=i[r]||function(){
(i[r].q=i[r].q||[]).push(arguments)},i[r].l=1*new Date();a=s.createElement(o),
m=s.getElementsByTagName(o)[0];a.async=1;a.src=g;m.parentNode.insertBefore(a,m)
})(window,document,'script','//www.google-analytics.com/analytics.js','ga');
ga('create', 'UA-45176973-1', 'openwetware.org');
ga('send', 'pageview');
</script> <html> <style type="text/css"> /*! fancyBox v2.1.5 fancyapps.com | fancyapps.com/fancybox/#license */ .fancybox-wrap, .fancybox-skin, .fancybox-outer, .fancybox-inner, .fancybox-image, .fancybox-wrap iframe, .fancybox-wrap object, .fancybox-nav, .fancybox-nav span, .fancybox-tmp { padding: 0; margin: 0; border: 0; outline: none; vertical-align: top; }
.fancybox-wrap { position: absolute; top: 0; left: 0; z-index: 8020; }
.fancybox-skin { position: relative; background: #f9f9f9; color: #444; text-shadow: none; -webkit-border-radius: 4px; -moz-border-radius: 4px; border-radius: 4px; }
.fancybox-opened { z-index: 8030; }
.fancybox-opened .fancybox-skin { -webkit-box-shadow: 0 10px 25px rgba(0, 0, 0, 0.5); -moz-box-shadow: 0 10px 25px rgba(0, 0, 0, 0.5); box-shadow: 0 10px 25px rgba(0, 0, 0, 0.5); }
.fancybox-outer, .fancybox-inner { position: relative; }
.fancybox-inner { overflow: hidden; }
.fancybox-type-iframe .fancybox-inner { -webkit-overflow-scrolling: touch; }
.fancybox-error { color: #444; font: 14px/20px "Helvetica Neue",Helvetica,Arial,sans-serif; margin: 0; padding: 15px; white-space: nowrap; }
.fancybox-image, .fancybox-iframe { display: block; width: 100%; height: 100%; }
.fancybox-image { max-width: 100%; max-height: 100%; }
- fancybox-loading, .fancybox-close, .fancybox-prev span, .fancybox-next span {
background-image: url('http://openwetware.org/images/5/55/Fancybox_sprite.png'); }
- fancybox-loading {
position: fixed; top: 50%; left: 50%; margin-top: -22px; margin-left: -22px; background-position: 0 -108px; opacity: 0.8; cursor: pointer; z-index: 8060; }
- fancybox-loading div {
width: 44px; height: 44px; background: url('http://openwetware.org/images/d/d0/Fancybox_loading.gif') center center no-repeat; }
.fancybox-close { position: absolute; top: -18px; right: -18px; width: 36px; height: 36px; cursor: pointer; z-index: 8040; }
.fancybox-nav { position: absolute; top: 0; width: 40%; height: 100%; cursor: pointer; text-decoration: none; background: transparent url('http://openwetware.org/images/c/c0/Blank.gif'); /* helps IE */ -webkit-tap-highlight-color: rgba(0,0,0,0); z-index: 8040; }
.fancybox-prev { left: 0; }
.fancybox-next { right: 0; }
.fancybox-nav span { position: absolute; top: 50%; width: 36px; height: 34px; margin-top: -18px; cursor: pointer; z-index: 8040; visibility: hidden; }
.fancybox-prev span { left: 10px; background-position: 0 -36px; }
.fancybox-next span { right: 10px; background-position: 0 -72px; }
.fancybox-nav:hover span { visibility: visible; }
.fancybox-tmp { position: absolute; top: -99999px; left: -99999px; visibility: hidden; max-width: 99999px; max-height: 99999px; overflow: visible !important; }
/* Overlay helper */
.fancybox-lock {
overflow: hidden !important; width: auto;
}
.fancybox-lock body {
overflow: hidden !important;
}
.fancybox-lock-test {
overflow-y: hidden !important;
}
.fancybox-overlay { position: absolute; top: 0; left: 0; overflow: hidden; display: none; z-index: 8010; background: url('http://openwetware.org/images/e/e0/Fancybox_overlay.png'); }
.fancybox-overlay-fixed { position: fixed; bottom: 0; right: 0; }
.fancybox-lock .fancybox-overlay { overflow: auto; overflow-y: scroll; }
/* Title helper */
.fancybox-title { visibility: hidden; font: normal 13px/20px "Helvetica Neue",Helvetica,Arial,sans-serif; position: relative; text-shadow: none; z-index: 8050; }
.fancybox-opened .fancybox-title { visibility: visible; }
.fancybox-title-float-wrap { position: absolute; bottom: 0; right: 50%; margin-bottom: -35px; z-index: 8050; text-align: center; }
.fancybox-title-float-wrap .child { display: inline-block; margin-right: -100%; padding: 2px 20px; background: transparent; /* Fallback for web browsers that doesn't support RGBa */ background: rgba(0, 0, 0, 0.8); -webkit-border-radius: 15px; -moz-border-radius: 15px; border-radius: 15px; text-shadow: 0 1px 2px #222; color: #FFF; font-weight: bold; line-height: 24px; white-space: nowrap; }
.fancybox-title-outside-wrap { position: relative; margin-top: 10px; color: #fff; }
.fancybox-title-inside-wrap { padding-top: 10px; }
.fancybox-title-over-wrap { position: absolute; bottom: 0; left: 0; color: #fff; padding: 10px; background: #000; background: rgba(0, 0, 0, .8); }
/*Retina graphics!*/ @media only screen and (-webkit-min-device-pixel-ratio: 1.5), only screen and (min--moz-device-pixel-ratio: 1.5), only screen and (min-device-pixel-ratio: 1.5){
#fancybox-loading, .fancybox-close, .fancybox-prev span, .fancybox-next span { background-image: url('http://openwetware.org/images/b/b8/Fancybox_sprite%402x.png'); background-size: 44px 152px; /*The size of the normal image, half the size of the hi-res image*/ }
#fancybox-loading div { background-image: url('http://openwetware.org/images/0/01/Fancybox_loading%402x.gif'); background-size: 24px 24px; /*The size of the normal image, half the size of the hi-res image*/ } }
- fancybox-buttons {
position: fixed; left: 0; width: 100%; z-index: 8050; }
- fancybox-buttons.top {
top: 10px; }
- fancybox-buttons.bottom {
bottom: 10px; }
- fancybox-buttons ul {
display: block; width: 166px; height: 30px; margin: 0 auto; padding: 0; list-style: none; border: 1px solid #111; border-radius: 3px; -webkit-box-shadow: inset 0 0 0 1px rgba(255,255,255,.05); -moz-box-shadow: inset 0 0 0 1px rgba(255,255,255,.05); box-shadow: inset 0 0 0 1px rgba(255,255,255,.05); background: rgb(50,50,50); background: -moz-linear-gradient(top, rgb(68,68,68) 0%, rgb(52,52,52) 50%, rgb(41,41,41) 50%, rgb(51,51,51) 100%); background: -webkit-gradient(linear, left top, left bottom, color-stop(0%,rgb(68,68,68)), color-stop(50%,rgb(52,52,52)), color-stop(50%,rgb(41,41,41)), color-stop(100%,rgb(51,51,51))); background: -webkit-linear-gradient(top, rgb(68,68,68) 0%,rgb(52,52,52) 50%,rgb(41,41,41) 50%,rgb(51,51,51) 100%); background: -o-linear-gradient(top, rgb(68,68,68) 0%,rgb(52,52,52) 50%,rgb(41,41,41) 50%,rgb(51,51,51) 100%); background: -ms-linear-gradient(top, rgb(68,68,68) 0%,rgb(52,52,52) 50%,rgb(41,41,41) 50%,rgb(51,51,51) 100%); background: linear-gradient(top, rgb(68,68,68) 0%,rgb(52,52,52) 50%,rgb(41,41,41) 50%,rgb(51,51,51) 100%); filter: progid:DXImageTransform.Microsoft.gradient( startColorstr='#444444', endColorstr='#222222',GradientType=0 ); }
- fancybox-buttons ul li {
float: left; margin: 0; padding: 0; }
- fancybox-buttons a {
display: block; width: 30px; height: 30px; text-indent: -9999px; background-color: transparent; background-image: url('fancybox_buttons.png'); background-repeat: no-repeat; outline: none; opacity: 0.8; }
- fancybox-buttons a:hover {
opacity: 1; }
- fancybox-buttons a.btnPrev {
background-position: 5px 0; }
- fancybox-buttons a.btnNext {
background-position: -33px 0; border-right: 1px solid #3e3e3e; }
- fancybox-buttons a.btnPlay {
background-position: 0 -30px; }
- fancybox-buttons a.btnPlayOn {
background-position: -30px -30px; }
- fancybox-buttons a.btnToggle {
background-position: 3px -60px; border-left: 1px solid #111; border-right: 1px solid #3e3e3e; width: 35px }
- fancybox-buttons a.btnToggleOn {
background-position: -27px -60px; }
- fancybox-buttons a.btnClose {
border-left: 1px solid #111; width: 35px; background-position: -56px 0px; }
- fancybox-buttons a.btnDisabled {
opacity : 0.4; cursor: default; }
- fancybox-thumbs {
position: fixed; left: 0; width: 100%; overflow: hidden; z-index: 8050; }
- fancybox-thumbs.bottom {
bottom: 2px; }
- fancybox-thumbs.top {
top: 2px; }
- fancybox-thumbs ul {
position: relative; list-style: none; margin: 0; padding: 0; }
- fancybox-thumbs ul li {
float: left; padding: 1px; opacity: 0.5; }
- fancybox-thumbs ul li.active {
opacity: 0.75; padding: 0; border: 1px solid #fff; }
- fancybox-thumbs ul li:hover {
opacity: 1; }
- fancybox-thumbs ul li a {
display: block; position: relative; overflow: hidden; border: 1px solid #222; background: #111; outline: none; }
- fancybox-thumbs ul li img {
display: block; position: relative; border: 0; padding: 0; max-width: none; } </style> </html>
<html>
<body>
<div id="subnav">
<ul id="navmenu" class="open">
<li><a href="#polymersome">Polymersome</a></li>
<li><a href="#origami">Origami channel</a></li>
<li><a href="#reaction">Internal reaction</a></li>
<li><a href="#nanoreactor">Nanoreactor</a></li>
<li><a href="#achievements">Achievements</a></li>
<li><a href="#future">Future work</a></li>
<li><a href="#applications">Applications</a></li>
<li><a href="#references">References</a></li>
</ul>
</div>
<section id="content" style="position: relative; top: -164px;">
<section id="motivation" class="project" data-stellar-background-ratio="0.5">
<h2>Motivation</h2> <div class="project_box" > <h3>'What I cannot create, I do not understand'</h3>
<p>This famous quote from Richard Feynman inspired our project. The initial idea of one physicist in our group was: <h3>Let's build a cell!</h3></p>
<p>So, of course, the biologists of our group tried to explain him, that a cell is far too complex and this is a crazy idea. If a biologist thinks of a cell, he sees an extremely complex structure, but the physicist just sees a simple black circle. So what is a cell, if you break it down to a simpler level?</p>
<p>A compartment separating a reactive center from the outside, combined with controlled means of transport in and out and some modifications on the outside. An intelligent, microscopically small reactor. And our idea of a </h3>Smart Nanoreactor</h3> was born.</p> </div>
<div class="figure_box"> <figure class="box " > <a class="fb" rel="motivation" href="http://openwetware.org/images/b/b8/Cell.png" title="A cell in biologist's perception (adapted from: http://www.aid-n.com/wp-content/uploads/2012/10/All-about-Human-Cells-Cell-in-Human.jpg)."> <img src="http://openwetware.org/images/4/4a/Cell_thumb.png" alt="" /> </a> <figcaption>Biologist's cell </figcaption> </figure>
<figure class="box " > <a class="fb" rel="motivation" href="http://openwetware.org/images/d/de/Physicists_cell.png" title="A cell in physicist's perception."> <img src="http://openwetware.org/images/c/ca/Physicists_cell_thumb.png" alt="" /> </a> <figcaption>Physicist's cell </figcaption> </figure> </div> <div class="clear"></div> </section>
<a class="anchor" id="polymersome"></a> <div class="interlude clearfix"><h2>Polymersome</h2></div>
<section id="vesicle" class="project" data-stellar-background-ratio="0.5"> <h2>Introduction</h2> <div class="project_box clearfix"> <p> First of all, we needed a compartment. In cells, the compartment is made out of a phospholipid bilayer membrane that forms the plasma membrane. But this being already nature's strategy, we wanted to try a different thing and decided to use block co-polymers to build the nanoreactor wall. </p> <!-- <p> The main goal of experiment was to synthesize stable and uniformly distributed polymersomes as vesicle for carrying the collagenase enzyme and also incorporating DNA origami channels for tunable delivery of cargo with respect to its size.</p> -->
<!-- PMOXA<sub>17</sub>-PDMS<sub>65</sub>-PMOXA<sub>17</sub> was choosen as our ABA tri-block polymer know for its high biocompatiblity and PEG<sub>45</sub>-b-P(DEAEMA<sub>10</sub>-stat-BMA<sub>65</sub>) as our AB di-block polymer which has an additional functional of being pH responsive and also property to encapsulate both hydrophilic and hydrophobic biological entities.</p> -->
<h3> What are polymers? </h3>
<p>Polymers consist of structural or repeating units of low molecular weight covalently connected to each other to give high molecular weight compounds. A polymer that consists of two different repeating units is referred to as a co-polymer (Odian, 1981). The covalent linkage of two intrinsically different homo polymers leads to the formation of block–copolymers. Block-copolymers consist of at least two, covalently bound, segments or blocks of different Homo polymers which lead to the formation of di-block or tri-block copolymers. For instance, a tri-block copolymer can have a general form A<sub>n</sub>-B<sub>m</sub>-C<sub>p</sub>, with A, B, C, being different monomer types constituting the different blocks. The subscripts n, m, and p stand for the degree of polymerization, i.e. the average number of each monomer present in each respective block (McGrath, 1981).</p>
<h3>Why using polymers for our Nanoreactor?</h3>
<p>There is growing interest in polymersomes due to their potential application in biotechnology and pharmacology (Blanazs et al., 2009) Currently, a great potential lies in their ability to mimic biological membranes by establishing aqueous three dimensional functional cavities. The stability of these membranes is higher than those of biological lipid-based vesicles. Thus, polymersomes consisting of amphiphilic block copolymers hold promise as alternative vesicles in biotechnological and biomedical applications (van Dongen et al., 2011).</p>
<p> Two important parameters of the membranes are their permeability and their stability. The advantage of synthetic block-copolymers as the building blocks of these nanoreactors is their higher stability over lipids due to the increased length, conformational freedom, and slower dynamics of the underlying polymers.</p>
<p>Other desired characteristics for the Nanoreactor the polymers can achieve are: <ul> <li> The shell has a hydrophilic, biocompatible low-protein-binding surface.</li> <li> The shell can act as a bio membrane for protein reconstitution.</li> <li> The nanovesicle can encapsulate hydrophilic substances.</li> <li> The shell material has enormous possibilities for molecular functionalization.</li> <ul></p> <p> The nanovesicles can thus be employed as specific targeted drug delivery systems (e.g. biotinylated vesicles), stimuli responsive nanocontainers (e.g. pH sensitive) or nanoreactors.</p>
<p>Towards the above goal, two polymers, an ABA tri-block polymer and an AB di-block polymer were investigated.</p> </div>
<div class="figure_box">
<figure class="box " >
<a class="fb" rel="introduction" href="http://openwetware.org/images/a/a9/Intro1.png" title="Comparison of a phospholipid bilayer of the plasma membrane with polymer walls.">
<img src="http://openwetware.org/images/5/5c/Intro1t.png" alt="" />
</a>
<figcaption>Polymers and Lipids</figcaption>
</figure>
<figure class="box " >
<a class="fb" rel="introduction" href="http://openwetware.org/images/2/2d/Polymersome.png" title="Comparison of polymersomes to other nanoparticle-based delivery technologies (source: vindicopharma.com/our-products).">
<img src="http://openwetware.org/images/f/f1/Polymersomet.png" alt="" />
</a>
<figcaption>Why polymersome?</figcaption>
</figure>
</div> <div class="clear"></div>
<h2>Tri-block polymer</h2> <div class="project_box"> <p>This ABA tri-block copolymer consists of a middle block of PDMS (poly (dimethyl siloxane)) and two PMOXA (poly (2-methyloxazoline)) side chains. These copolymers self-assemble in aqueous solution to form highly stable, closed vesicles with a controlled diameter of typically less than 200 nm.</p>
<p>Tri-block polymersomes were prepared by film rehydration using PBS buffer (pH 7.4) as rehydrating solvent. The polymersomes were observed using cryo-SEM and cryo-TEM, and the particle size was characterized by DLS (Dynamic light scattering).</p>
<p>Cryo-TEM images clearly depict lamellar and large vesicle formation, which is further confirmed by DLS showing an average diameter of ~950nm and a broad particle size distribution. These are not as desired, so these polymersomes were further subjected to an extrusion process using Ultra-filtration equipment.</p>
<p> After extrusion, the samples were observed under cryo-TEM again and their size distribution was measured by DLS. Ultra-filtration process extruded larger vesicles and lamellar structure which is well seen in TEM images. In the DLS measurement, the peak of diameter ranging from 226 to 238 nm is better defined and corresponds to 50% of the total polymersomes.</p> </div>
<div class="figure_box"> <figure class="box " > <a class="fb" rel="polymer" href="http://openwetware.org/images/d/d7/PMOXA_PDMS_PMOXA.png" title="Tri-block Copolymer: ABA Tri-block Copolymer consists of PDMS (poly(dimethyl siloxane)) middle block and two PMOXA (poly(2-methyloxazoline)) side chains (source: Grumelard 2004)."> <img src="http://openwetware.org/images/e/eb/ABApolymer.png" alt="" /> </a> <figcaption>Tri-block polymer </figcaption> </figure>
<figure class="box " > <a class="fb" rel="polymer" href="http://openwetware.org/images/b/b1/Pbs_unfiltered_lamellar_structures.png" title="Unfiltered tri-block polymers: Cryo-TEM image of unfiltered tri-block polymersome indicating large lamellar structure."> <img src="http://openwetware.org/images/9/91/Pbs_poly1_026_thumbs_final_thumb.png" alt="" /> </a> <figcaption>Unfiltered tri-block polymers</figcaption> </figure>
<figure class="box " > <a class="fb" rel="polymer" href="http://openwetware.org/images/f/f8/Collage.png" title="Filtrated polymersome: Cryo-TEM image of filtered tri-block polymersome using ultra-filtration equipment. The scale bar is 200nm."> <img src="http://openwetware.org/images/a/a4/Poly_1_thumb_final_thumb.png" alt="" /> </a> <figcaption>Filtrated polymersome</figcaption> </figure>
<figure class="box " >
<a class="fb" rel="polymer" href="http://openwetware.org/images/8/8d/DLS_final.png" title="DLS measurement for tri-block polymersomes. After filtration, the DLS measurement shows a more defined peak for the polymersomes of 200 nm diameter, which correspond to 50 % of the total polymersomes.">
<img src="http://openwetware.org/images/b/b7/DLS_thumb.png" alt="" />
</a>
<figcaption>DLS measurement</figcaption>
</figure>
</div>
<div class="clear"></div>
<h2>Di-block polymer</h2> <div class="project_box" > <p>We were also interested in thermo-responsive block copolymer, poly(2-cinnamoylethyl methacrylate)-poly(N-isopropylacryl amide), pH-sensitive and photo-cross-linkable block copolymer. They can function as on/off switches depending on the external pH stimulus due to the pH sensitivity of their cross-linked membrane.</p>
<p>The block copolymers undergo spontaneous self-assembly into the desired polymersome structures by raising the pH of the aqueous solution from 3 to 10. To preserve the vesicular shape of the polymersomes under varying conditions, cross-linking of their membranes is highly desired.</p>
<p>These di-block polymers self-assemble into polymersomes of uniform size of 100-120 nm and later expand till 140 nm as the pH is lowered from alkaline condition to acidic state. Thus, they act as pH responsive polymersomes which can tune its release of biological entities based on environmental pH condition. Such property makes them demandable in field of drug therapy specifically in treating cancer.</p>
<p>The formation of the polymersomes was analysed using cryo-TEM and the particle size distribution was measured with DLS. In this case, the distribution is very uniform with a peak at 140 nm.</p>
<p>The pH responsive nature of di-block polymer was investigated by the release profile of Doxorubicin, an anti-cancerogenic agent which showed slow release at pH 7.4 and faster release at pH 5. This is a proof that the polymersomes were effectively cross-linked by UV.</p> </div>
<div class="figure_box">
<figure class="box " >
<a class="fb" rel="polymer2" href="http://openwetware.org/images/7/71/Di_block_polymer.png" title="Di-block polymer: Chemical structure of the pH-responsive di-block polymer. The porosity of the walls changes according to pH (source: Yassin et al., 2012). ">
<img src="http://openwetware.org/images/a/a5/Di_block_polymer_thumb.png" alt="" />
</a>
<figcaption>Di-block polymer </figcaption>
</figure>
<figure class="box " > <a class="fb" rel="polymer2" href="http://openwetware.org/images/2/2c/Poly2_011.png" title="Di-block polymersomes: Cryo-TEM image of Di-block polymersomes showing uniformly distributed polymersomes."> <img src="http://openwetware.org/images/e/ec/Poly2_thumb_thumb.png" alt="" /> </a> <figcaption>Cryo-TEM </figcaption> </figure>
<figure class="box " > <a class="fb" rel="polymer2" href="http://openwetware.org/images/f/f9/DLS_poly_2.png" title="DLS measurement: diameter distribution of the particles shows a peak at 140 nm."> <img src="http://openwetware.org/images/6/6b/DLS_poly_2_final_thumb.png" alt="" /> </a> <figcaption>DLS measurement</figcaption> </figure>
<figure class="box " > <a class="fb" rel="polymer2" href="http://openwetware.org/images/9/9c/Release_profile.png" title="Release profile: Release profile of Doxorubicin by di-block polymersomes at pH 5 and pH 7."> <img src="http://openwetware.org/images/9/9a/Release_poly_thumb.png" alt="" /> </a> <figcaption>Release profile</figcaption> </figure> </div> <div class="clear"></div> </section>
<a class="anchor" id="origami"></a> <div class="interlude"><h2>DNA Origami channel</h2></div>
<section id="origami" class="project" data-stellar-background-ratio="0.5"> <h2>Introduction</h2> <div class="project_box"> <p> To allow transport inside and outside the smart nanoreactor, a channel is needed. Instead of using a protein channel, we decided to use a DNA origami channel, because it is more robust, and we can easily design it according to the characteristics of the polymer wall.</p> <p> <h3>What is DNA Origami?</h3> </p> <p>In DNA origami a circular ssDNA molecule of several thousand base pairs, called scaffold, is folded with the help of short ssDNA strands, the staple strands. The staple strands bring together two different zones of the scaffold, and by varying their sequences different 2D or 3D shapes can be achieved.</p>
<p>DNA origami has unique properties, such as an addressable surface, which enables selective functionalization with biomolecules and nanomaterials. The origami can also be combined with top-down nanotechnology, such as placement on a fabricated substrate. The technology can also be used in single-molecule imaging, where FRET pairs or fluorophores can be constructed on designed DNA origami structures. Furthermore, DNA origami can be easily combined with other DNA nanodevices such as DNAzymes, DNA beacons or DNA walkers acting on DNA origami. Hence, DNA origami technology has practical potential in various research fields.</p> </div> <div class="figure_box"> <figure class="box " > <a class="fb" rel="origami" href="http://openwetware.org/images/b/b2/Origami_explanation.png" title="DNA origami: In DNA origami, the scaffold - represented in black - is folded by the staple strands - represented in different colours - into a 2D or 3D shape. Adapted from: Rothemund et al, Nature 2006"> <img src="http://openwetware.org/images/8/8c/Origami_explanation_thumb.png" alt="" /> </a> <figcaption> DNA origami </figcaption> </figure> </div> <div class="clear"></div> <h2>Design</h2> <div class="project_box" > <h3>Requirements</h3> <p> The origami channel was designed to be properly integrated into the polymersome wall. For this, the dimensions were restricted to the thickness of the wall, as well as to the length of the scaffold. Furthermore, the dimensions of the porus are determined by the dimensions of the substrate and the enzyme: it has to be big enough so the substrate and products of the reaction can easily go through, but small enough so the enzyme stays encapsulated inside the polymersome. The final dimensions are the following: <ul> <li>Length: 34 nm</li> <li>Diameter: 25 nm</li> <li>Pore diameter: 6 nm</li> </ul> </p>
<h3>Scaffold and staple types</h3>
<p>For the design, the caDNAno program was used (see http://cadnano.org/), and the stability of the structure was tested with CanDo (see http://cando-dna-origami.org/).</p> <p>The scaffold used in this design is 7560 bp long, derived from the M13p18 Escherichia coli virus. Its sequence was taken from the cadnano design program. Bending it into a structure with a 78 helix bundles, the desired length is achieved. The channel contains four types of staples:</p> <p><ul> <li>core: 118 staples give stability to the whole structure.</li> <li>edge: 39 staples give stability to the edges.</li> <li>helpers: 12 staples have a 5' protruding end with 5 adenines, which should help the anchors point to the right directions.</li> <li>anchors: 12 staples have a 3' protruding end of 21 nucleotides (7 nm) which will allow the binding of the oligonucleotides that carry the hydrophobic modifications. Between the corresponding complementary sequence to the scaffold and the protrusion, 5 thymine nucleotides serve as spacer and hybridize with the 5 adenines present in the helpers. </li> <li>no_hang: can be used instead of the anchors to generate a channel without any anchors sticking out of the channel walls. They can serve as a negative control when introducing the hydrophobically modified oligonucleotides.</li> </ul></p> <p>Their sequences and also the map of their distribution in the scaffold can be donwnloaded from the lab book.</p> </div> <div class="figure_box"> <figure class="box " > <a class="fb" rel="origami" href="http://openwetware.org/images/c/c0/DNA_channel.gif" title="DNA origami channel: 3D model of our DNA origami channel design."> <img src="http://openwetware.org/images/5/55/DNA_channel_thumb.gif" alt="" /> </a> <figcaption>DNA origami channel</figcaption> </figure>
<figure class="box " > <a class="fb" rel="origami" href="http://openwetware.org/images/a/af/Front_view.png" title="Front view: Front view of the DNA origami channel generated with the caDNAno program."> <img src="http://openwetware.org/images/1/1c/Front_view_thumb.png" alt="" /> </a> <figcaption>Front view </figcaption> </figure>
<figure class="box " > <a class="fb" rel="origami" href="http://openwetware.org/images/d/da/Lateral_view.png" title="Lateral view: Lateral view of the DNA origami channel generated with the caDNAno program."> <img src="http://openwetware.org/images/8/81/Lateral_view_thumb.png" alt="" /> </a> <figcaption>Lateral view </figcaption> </figure>
</div>
<div class="clear"></div> <h2>Folding and characterization</h2> <div class="project_box"> <p> The mixture of scaffold and different staples was subjected to a thermal annealing ramp, that allowed the folding. In an initial experiment, different concentrations of Magnesium chloride in the folding buffer were tested, and the resulting structures were analized by atomic force microscopy and electrophoresis to determine the best concentration. The range 10 to 14 mM MgCl<sub>2</sub> were the best, so we chose the 12 mM concentration for further experiments.</p> <p>In the AFM images the height and width were measured, obtaining the approximate values of 7 and 60 nm respectively. The height appears to be much lower than the expected, due to the high absorption of the channel to the substrate - mica - used for imaging. Mica is negatively charged, and so the Magnesium chloride in the samples interacts with the surface creating a positively charged substrate. Thus, the DNA origami channel can interact tightly with the surface, modifying its structure. The width appears much bigger, which can be due to the tip-sample convolution.</p> </div>
<div class="figure_box">
<figure class="box " >
<a class="fb" rel="fold" href="http://openwetware.org/images/4/42/Electro_magchl.png" title="Electrophoresis: Electrophoresis of the channels after folding and filtration using the stated MgCl<sub>2</sub> concentrations in the folding buffer.">
<img src="http://openwetware.org/images/2/21/Electro_magchl_thumb.png" alt="" />
</a>
<figcaption>Electrophoresis </figcaption>
</figure>
<figure class="box " > <a class="fb" rel="fold" href="http://openwetware.org/images/3/3f/AFM_magchl.png" title="AFM folding analysis: Corroboration by AFM imaging of the folded or unfolded states of the DNA origami channels after filtration, using folding buffers containing different concentration of MgCl<sub>2</sub>"> <img src="http://openwetware.org/images/c/c7/AFM_magchl_thumb.png" alt="" /> </a> <figcaption>AFM folding analysis </figcaption> </figure>
<figure class="box " > <a class="fb" rel="fold" href="http://openwetware.org/images/9/99/AFM_quantification.png" title="AFM measurements: AFM imaging of the folded DNA origami channels after filtration. The graph is the superposition of the height profiles from 20 different structures. From these, the height and width, shown in the table, were calculated. </figcaption> </figure>"> <img src="http://openwetware.org/images/0/0b/AFM_quantification_thumb.png" alt="" /> </a> <figcaption> AFM measurements </figcaption> </figure> </div>
<div class="clear"></div>
<h2>TEM analysis</h2>
<div class="project_box">
<p> The 3D structure was then corroborated using TEM and cryo-TEM.</p>
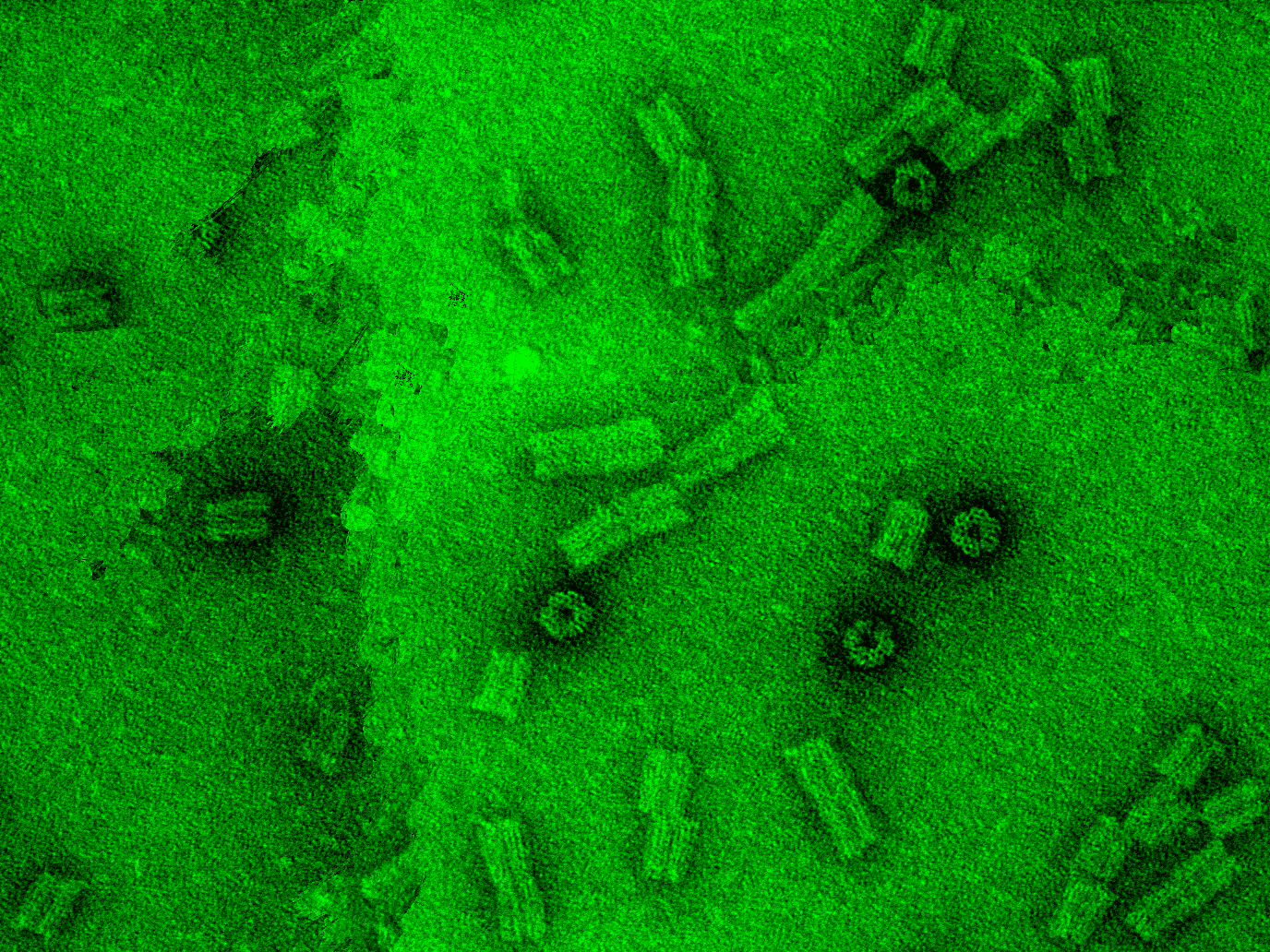
<p>For the TEM images, the diameter and length of 20 different structures were measured. In this case, the dimensions of the channel and also its shape correspond accurately to the design. </p> </div> <div class="figure_box"> <figure class="box " > <a class="fb" rel="tem" href="http://openwetware.org/images/d/df/TEM_channel_overview_final.png" title="TEM origami channel: Typical TEM image of the DNA origami channels after negative staining with uranyl acetate 2%."> <img src="http://openwetware.org/images/6/62/TEM_channel_overview_final_thumb.png" alt="" /> </a> <figcaption>TEM origami channel </figcaption> </figure>
<figure class="box " > <a class="fb" rel="tem" href="http://openwetware.org/images/2/27/TEM_channel_amplification_final_white.png" title="TEM origami channel: High magnification of TEM images of different DNA origami channels after negative staining with uranyl acetate. The scale bar is 20nm."> <img src="http://openwetware.org/images/5/5f/TEM_channel_amplification_final_thumb.png" alt="" /> </a> <figcaption>TEM origami channel </figcaption> </figure>
<figure class="box " > <a class="fb" rel="tem" href="http://openwetware.org/images/f/fd/Cryo_final.png" title="Cryo-TEM origami channel: cryo-TEM of the DNA origami channel."> <img src="http://openwetware.org/images/1/18/Cryo_TEM_final_thumb.png" alt="" /> </a> <figcaption>Cryo-TEM origami channel </figcaption> </figure>
<figure class="box " > <a class="fb" rel="tem" href="http://openwetware.org/images/7/7d/Measurement_final.png" title="Measurement of the dimensions of the 20 channels from TEM images."> <img src="http://openwetware.org/images/1/19/Measurement_finalt.png" alt="" /> </a> <figcaption>TEM measurements </figcaption> </figure> </div>
<div class="clear"></div>
<h2>Hydrophobic modifications</h2>
<div class="project_box">
<p>For the channel to be incorporated inside the polymersome membrane spontaneously, a hydrophobic modification was introduced to a short oligonucleotide that hybridizes with the anchors. Two different types of hydrophobic modifications were performed.</p>
<h3>5' Palmitate and arachidic acid modification</h3> <p>An amino group was introduced to the 5' end of the oligonucleotides, that afterwards reacts with the carboxylic group of the molecule containing the hydrophobic fatty acids palmitate and arachidate. The reaction used is an N-acylation, which requires some activations. More details of the reaction and reagents used can be found in the Lab book.</p>
<p>The modified oligos were purified with HPLC and the purity of the product was analized by mass spectrometry.</p>
<h3>3' Cholesterol modification</h3>
<p>Oligonucleotides for 3' cholesterol modification were synthesized on a solid support already carrying the cholesterol molecules. In this case the sequence was shorter for the modification to be slightly separated from the walls of the channel. The modified oligos were purified with HPLC and the purity of the product was analized with mass spectrometry.</p>
<h3>Incorporation of the hydrophobically-modified oligonucleotides into the origami channel</h3> <p> The success of the incorporation of the hydrophobically modified oligos into the DNA origami channel was analyzed by electrophoresis.</p> <p/> The DNA origami channel was folded with the presence of no_hang staples, so a structure with no anchors was produced. This channel without anchors was put into contact with the oligonucleotide complementary to the anchors without modification, and also with the oligonucleotide with two different types of hydrophobic modifications. This served as a negative control as no difference in the electrophoretic mobility should be observed.</p> <p> In parallel, the DNA origami channel with the anchor staples was produced in the present of the oligonucleotide complementary to them without the hydrophobic modifications, and the same oligonucleotide with two different types of modifications.</p> <p> In the electrophoresis gel a shift can be observed between the electrophoresis mobility of the channels which don't contain the anchors and the ones which contain them. This is because the shape of the structure is affected by these sticking out DNA strands in a way that they cannot run as fast in the gel. Moreover, when introducing the oligonucleotides modified with 5' palmitic and arachidic acid that hybridize with the anchors, a further shift is observed, indicating that this modification is correctly incorporated into the channel. As for the 3' cholesterol modification, no shift is observed, but this can be due to the small volume of the cholesterol molecules in respect to the channel and their position being closer to the channel walls. In both modifications, though, a remaining of the sample inside the walls can indicate a certain level of formation of big complexes unable to enter the gel due to aggregation of the channel through their hydrophobic regions, specially in the 5' palmitic and arachidic acid modified channel. This is a good indicator that the hydrophobic modifications were properly incorporated onto the DNA origami channel. </p> </div>
<div class="figure_box"> <figure class="box " > <a class="fb" rel="mod" href="http://openwetware.org/images/0/0c/Purif_palm_arach.png" title="HPLC purification and mass spectrometry analysis of the oligonucleotides modified with palmitate and arachidic acid. A. HPLC chromatogram of the first HPLC purification round. The peak at almost 26 minutes corresponds with the expected elution time of the product, but being so low an elution with methanol was required. B. HPLC chromatogram of the second HPLC purification round, where the product was eluted with methanol. The fraction number 21 was used for the further experiments. C. Mass spectrometry chromatogram of the fraction number 21, the peak corresponds to the product. D. Mass spectrometry spectrum of the amplified peak, the molecular weight of the oligonucleotide with the correct hydrophobic modification is 7270 Da. The highest peak with an m/z of 1815.79 on the right corresponds with the species with 4 positive charges. Other peaks correspond to different number of charges."> <img src="http://openwetware.org/images/9/95/Purif_palm_arach_thumb.png" alt="" /> </a> <figcaption>5' modification purification</figcaption> </figure>
<figure class="box " > <a class="fb" rel="mod" href="http://openwetware.org/images/a/a3/Purif_chol.png" title="HPLC purification and Mass spectrometry analysis of the oligonucleotides modified with cholesterol. A. HPLC chromatogram. The peak eluting at 10,5 minutes corresponds to the cholesterol-modified oligo. The fraction nr. 36 was used for further experiments. B. Mass spectrometry chromatogram of the fraction number 36, the peak corresponds to the product. D. Mass spectrometry spectrum of the amplified peak, the molecular weight of the oligonucleotide with the cholesterol modification is 5387 Da. The highest peak wit m/z of 1793.86 on the right corresponds with the species with 3 positive charges. Other peaks correspond to different number of charges."> <img src="http://openwetware.org/images/6/6c/Purif_chol_thumb.png" alt="" /> </a> <figcaption>3' modification purification</figcaption> </figure>
<figure class="box " >
<a class="fb" rel="mod" href="http://openwetware.org/images/8/88/Electro_hydrophobic_final.png" title="Electrophoresis to analyze if the hydrophobically modified oligonucleotides bind correctly to the anchors in the origami channel.">
<img src="http://openwetware.org/images/1/15/Electro_hydrophobic_final_thumb.png" alt="" />
</a>
<figcaption>Electrophoresis </figcaption>
</figure>
</div>
<div class="clear"></div>
</section>
<a class="anchor" id="reaction"></a> <div class="interlude clearfix"><h2>Internal reaction</h2></div>
<section id="reaction" class="project" data-stellar-background-ratio="0.5">
<h2>Introduction</h2>
<div class="project_box" > <p>The aim of our project is to create a nanoreactor. Inspired by how cell metabolism is organized in specific cellular compartments, we decided to design a system which is capable of hosting enzymatic reactions. Inner compartmentalization allows fragile processes to remain protected against undesired influences, such as proteolytic or microbial degradation and also one could expect enhanced reaction probabilities and efficiencies due to spatial confinement or immobilization of enzymes.</p> </div> <div class="clear"></div>
<h2>Choice of reaction</h2>
<div class="project_box" > <p>There are several requirements for the enzyme which can be entrapped. The enzymes should be large enough so that they are not able to escape, especially if there are channels or pores formed in the system. Another requirement is that the reaction is easy to detect. Examples include enzymes which are able to produce fluorescent or chromogenic products, such as myoglobin, Candida Antarctica lipase B, glucose oxides, horse radish peroxidase, urease and α-chymotrypsin. </p> <p>The protein of our choice was collagenase. Collagenases are endopeptidases that degrade the helical regions in native collagen preferentially at the Y-Gly bond in the sequence Pro-Y-Gly-Pro, where Y is most frequently a neutral amino acid. This protein is very abundant in many species and it is easy to obtain from the bacteria Clostridium hystoliticum. The isolated enzyme is composed of 7 different proteases ranging in molecular weight from 68-130 kDa, thus the protein is big enough and there is no possibility to pass through the channel. Furthermore, the reaction can be detected easily with spectrophotometric assay.</p> </div>
<div class="clear"></div>
<h2>Fluorescent labeling</h2>
<div class="project_box" > <p>The protein was conjugated to amine-reactive fluorescent dye in order to prove its entrapment inside the polymersomes. TAMRA NHS was used to perform the fluorescent labeling. TAMRA (Tetramethylrhodamine) belongs to the group of the long-wavelength rhodamines with excitation maximum at 555nm and emission maximum at 580 nm.</p>
<p>The degree of labelling with TAMRA dye was estimated to be 18 dye molecules per enzyme.</p>
</div> <div class="clear"></div>
<h2>Enzyme reaction assay</h2>
<div class="project_box" > <p>The enzymatic assay for determining the rate of the reaction was carried out using the peptide FALGPA, standing for N-(3-[2-furyl]acryloyl)-Leu-Gly-Pro-Ala. There is sufficient structural similarity between the 2-furanacryoyl group and Pro for FALGPA to be an excellent substrate for collagenase.</p>
<p>The molecular weight of the substrate is 476 Da. The maximum molecular weight of the molecules that can go through the polymer walls being 400 Da, the substrate will not be able to penetrate the polymer walls, but only to enter the nanoreactor through the channel. Furthermore, the substrate being uncharged, the negative charge of the DNA won't interfere with its transport.</p>
<p>This peptide has absorbance at 345 nm and the reaction is monitored through continuous spectrophotometric measurement at this wavelength. The assay is made possible by the blue shift in the near-ultraviolet absorption band of the furanacryloyl peptide when the peptide bond between the first and second residues is hydrolyzed, which means that after cleavage of the peptide, the absorbance at 345nm begins to decrease. The enzyme activity before and after conjugation to the dye is characterized through this assay. </p>
<p>The conjugation to the fluorescent dye leads to a slight decrease in the activity of the enzyme. The FALGPA units are calculated, taking into account the slope of the linear fit. The unmodified enzyme is estimated to have 3 FALGPA units/ml and the conjugated one – 2.6 FALGPA units/ml.</p>
<p>Since the polymersomes can be source of light scattering events, UPLC measurement of the reaction products was applied for better accuracy of our experimental results.</p> </div> <div class="figure_box"> <figure class="box " > <a class="fb" rel="rct" href="http://openwetware.org/images/3/34/IR_substrate.png" title="The substrate for the enzymatic reaction: N-(3-[2-FURYL]ACROYL) -LEU-GLY-PRO-ALA"> <img src="http://openwetware.org/images/f/f6/IR_substrate_thumb.png" alt="" /> </a> <figcaption>Substrate</figcaption> </figure>
<figure class="box " > <a class="fb" rel="rct" href="http://openwetware.org/images/a/a6/Nm_figure2.png" title="Reaction assay with unmodified enzyme. Continuous sprectrophotometric measurement at a wavelength of 345nm. The recorded decrease in absorbance is fitted using linear regression."> <img src="http://openwetware.org/images/d/d4/Figure2_thumb.png" alt="" /> </a> <figcaption>Unmodified enzyme</figcaption> </figure>
<figure class="box " > <a class="fb" rel="rct" href="http://openwetware.org/images/9/9a/Nm_figure3.png" title="Reaction assay with conjugated enzyme. Continuous spectrophotometric measurement at a wavelength of 345nm. The recorded decrease in absorbance is fitted using linear regression."> <img src="http://openwetware.org/images/1/19/Figure3_thumb.png" alt="" /> </a> <figcaption>Conjugated enzyme</figcaption> </figure>
<figure class="box " >
<a class="fb" rel="rct" href="http://openwetware.org/images/7/70/Nm_figure4.png" title="Comparison between the reaction rate of unmodified and conjugated enzyme.">
<img src="http://openwetware.org/images/0/0e/Figure4_thumb.png" alt="" />
</a>
<figcaption>Unmodified and conjugated enzyme</figcaption>
</figure>
<figure class="box " > <a class="fb" rel="rct" href="http://openwetware.org/images/0/0b/UPLC_enzyme.PNG" title="UPLC measurements. The substrate and the products can be distinguished by their different retention times. A. Chromatogram of the UPLC measurement of the substrate. B. Chromatogram of the products obtained with the conjugated enzyme reaction. C. Chromatogram of the products obtained with the unmodified enzyme reaction. It can be observed that in the case of the conjugated enzyme both peaks are present, which is due to the decrease in enzymatic activity after conjugation to the fluorescent dye."> <img src="http://openwetware.org/images/c/cf/UPLC_thumb.png" alt="" /> </a> <figcaption>UPLC measurements</figcaption> </figure> </div> <div class="clear"></div>
</section>
<a class="anchor" id="nanoreactor"></a>
<div class="interlude clearfix"><h2>Smart nanoreactor</h2></div>
<section id="nanoreactor" class="project" data-stellar-background-ratio="0.5">
<h2>Incorporation of the channel </h2>
<div class="project_box"> <p>The DNA origami channel was incorporated during the polymersome preparation and was characterized using TEM and Cryo-TEM.</p>
<p>For the tri-block copolymer, the polymersome formation was not affected by the addition of the DNA origami channel, and the DLS analysis confirmed that the polymersome size distribution was the same. In some of the polymersomes, particles across the membrane were observed. This indicates that the origami channels were successfully incorporated.</p>
<p>To further confirm the incorporation of the channel in the wall, gold nanoparticles can be attached to the DNA channel. This will make them more distinguishable in the TEM and cryo-TEM analysis. This is planned as a follow-up experiment.</p>
<p>The functionality of the channel was tested through a dye release assay. As a further confirmation of the incorporation of the channel in the membrane, and its functionality, we performed a dye release assay.</p>
<p>For the di-block copolymer, the polymersome formation seemed to be affected by the presence of the DNA origami channel. The polymersome membrane surface became rough and also its stability decreased.The polymersomes had a smaller diameter than previously observed and were present in a smaller amount. This difficulted the observation of the DNA channel incorporation.</p>
</div> <div class="figure_box"> <figure class="box " > <a class="fb" rel="channel" href="http://openwetware.org/images/7/7d/Incorp_channel_col_arrows.png" title="Cryo-TEM images of incorporated DNA channel with cholesterol modification in Tri-block polymersomes (scale bar = 100nm). The green arrows indicate the successful incorporation of the cholesterol-modified DNA channels."> <img src="http://openwetware.org/images/8/84/Incorp_channel_col_thumb.png" alt="" /> </a> <figcaption>Tri-block DNA-chol</figcaption> </figure>
<figure class="box " > <a class="fb" rel="channel" href="http://openwetware.org/images/1/1f/Incorp_2_scale_bar.png" title="Cryo-TEM images of incorporated DNA channel with palmitate and archidic modification in Tri-block polymersomes. The green arrows indicate the successful incorporation of the palmitate and arachidic acid-modified DNA channels."> <img src="http://openwetware.org/images/4/41/Collage_thumb.png" alt="" /> </a> <figcaption>Tri-block DNA-P+A</figcaption> </figure>
<figure class="box " > <a class="fb" rel="channel" href="http://openwetware.org/images/3/3d/Nm_collage_3.png" title="Cryo-TEM images of Di-block polymersomes in presence of DNA channel with cholesterol modification (scale bar = 200nm). The number of polymersomes were smaller than previously observed and also present in a lower amount."> <img src="http://openwetware.org/images/b/b9/Dbdnachol.png" alt="" /> </a> <figcaption>Di-block DNA-chol </figcaption> </figure> <figure class="box " > <a class="fb" rel="channel" href="http://openwetware.org/images/a/a9/Nm_collage_4.png" title="Cryo-TEM images of di-block polymersomes in presence of DNA channel with palmitate and archidic modification (scale bar = 200nm). The number of polymersomes were smaller than previously observed and also distorted by morphology."> <img src="http://openwetware.org/images/d/db/Dbdnapa.png" alt="" /> </a> <figcaption>Di-block DNA-chol </figcaption> </figure> </div> <div class="clear"></div>
<h2>Dye release assay</h2>
<div class="project_box"> <p>In order to confirm the functionality of the origami channels and their successful integration into the polymersome walls, a dye release experiment was performed. The fluorescein dye was encapsulated into the polymersomes in presence and absence of the channels. Fluorescein molecular weight is 332.29 Da and it can pass through the polymer walls, although with some difficulties. Thus, the incorporation of the channel should facilitate the exit of the dye. All the samples were placed inside dialysis bags and the fluorescein concentration outside the polymersomes was measured by UV/vis spectroscopy.</p>
<p>As a positive control, the release of fluorescein from the dialysis bag without polymersomes was also performed.</p>
<p> Tri-block polymersomes show around 80% of release within 24h whereas tri-block polymersomes with DNA channels exhibit complete release of dye. This is a strong hint of the successful incorporation of DNA channel within the tri-block polymersome walls, and that those are functional and allow the fluorescein to go through.</p>
<p> Di-block polymersomes with and without origami channels have similar release profile. The release of dye from the di-block polymersome was less than 50% which proves association of dye with polymer. It is in agreement with confocal LSM images presenting strong adhesion of the dye molecules with di-block polymer which may inhibit polymersome formation. In this case, thus, the dye-release assay cannot be used to prove the incorporation of the channels due to the interaction of the dye with the polymersome walls.</p>
<p>After 24h, the samples were also imaged by C-LSM (Confocal Laser Scanning Microscopy).</p>
<p> Tri-block Polymersomes without channels clearly trapped the dye molecules which could be observed in C-LSM images, whereas the polymersome with DNA channels released most of the dye and residual polymers containing dye could only be imaged with 10x higher laser power and increased gain. This shows that in the latter case, almost all the dye could go outside of the polymersomes.</p>
<p> Di-block polymersomes were highly destabilized in presence of dye. Only a few polymersomes encapsulated dye molecules similar to tri-block polymersome. Most polymersome were in disrupted state and strongly adhering fluorescence dye molecules. This result is in agreement with cryo-TEM images were we observed the similar structure of polymersome. </p> </div>
<figure class="box " > <a class="fb" rel="Dye assay" href="http://openwetware.org/images/f/fa/Pol1_dialysis.PNG" title="Dye Release profile for Tri-block Polymersomes"> <img src="http://openwetware.org/images/9/98/Pol1_dialysis_thumb.png" alt="" /> </a> <figcaption>Dye assay with Tri-block polymersome</figcaption> </figure>
<figure class="box " > <a class="fb" rel="Dye assay" href="http://openwetware.org/images/f/f6/Pol2_dialysis.PNG" title="Dye Release profile for Di-block Polymersomes"> <img src="http://openwetware.org/images/c/c0/Pol2_dialysis_thumb.png" alt="" /> </a> <figcaption>Dye assay with Di-block polymersome</figcaption> </figure>
<figure class="box " > <a class="fb" rel="Dye assay" href="http://openwetware.org/images/2/2b/Confocal_dye_triblock.png" title="Confocal-LSM image for Tri-block Polymersome after 24hr Dialysis. On the left, the control of the polymersomes without channel shows dye still inside the polymersomes, whereas in the presence of both cholesterol-modified or palmitate-arachidic-acid- modified channels 10 times more laser power was needed to see at least some remainigs of the dye inside. "> <img src="http://openwetware.org/images/a/a1/Pol1_clsm.png" alt="" /> </a> <figcaption>C-LSM of Tri-block polymersome</figcaption> </figure>
<figure class="box " > <a class="fb" rel="Dye assay" href="http://openwetware.org/images/f/fa/Confocal_dye_diblock.png" title="Confocal-LSM image for Di-block Polymersome after 24hr Dialysis. On the left, the control of the dye encapsulated inside the polymersoes without DNA channels, which presents less incorporation than with the Tri-block polymersomes. With DNA channel modified with cholesterol and palmitate-archidic similar images were obtained."> <img src="http://openwetware.org/images/2/2c/Pol2_clsm.png" alt="" /> </a> <figcaption>C-LSM of Di-block polymersome</figcaption> </figure>
<div class="clear"></div>
<h2>Encapsulation of the enzyme</h2>
<div class="project_box" > <p>The collagenase conjugated with the TAMRA fluorescent dye was encapsulated in both polymersomes. Dialysis was performed to eliminate the non-encapsulated enzyme. The encapsulation efficiency was calculated as the concentration after dialysis divided by the concentration before dialysis. For the Tri-block polymersomes, it was of around 70%, whereas for Di-block polymersomes, it resulted to be of around 30%.</p>
<p>The samples were then imaged by C-LSM. Particles that correspond to the polymersomes with the encapsulated enzyme could be observed in both Tri-block and Di-block polymers.</p>
<p>In order to prove that after dialysis all the free enzyme is removed, the polymerosmes were incubated with substrate solution and UPLC measurements were taken at different intervals of time. Even after 16 h of incubation, no products were detected, meaning that no free enzyme is present in the solution and that the enzyme is successfully encapsulated inside the polymersomes.</p>
</div>
<div class="figure_box">
<figure class="box " > <a class="fb" rel="Enzyme" href="http://openwetware.org/images/2/22/Nm_collage_5.png" title="C-LSM images of enzyme encapsulation in Tri-block polymersome under dynamic motion after dialysis"> <img src="http://openwetware.org/images/2/27/Clsmpoly1enzymet.png" alt="" /> </a> <figcaption>Tri-block enzyme encapsulation</figcaption> </figure>
<figure class="box " > <a class="fb" rel="Enzyme" href="http://openwetware.org/images/f/f6/Nm_collage_6.png" title="C-LSM image of enzyme encapsulation in Di-block polymersome under dynamic motion after dialysis"> <img src="http://openwetware.org/images/7/7e/Collage_6t.png" alt="" /> </a> <figcaption>Di-block enzyme encapsulation</figcaption> </figure>
<figure class="box " > <a class="fb" rel="Enzyme" href="http://openwetware.org/images/e/e4/Nm_uplc1.png" title="UPLC measurement to check for the presence of free enzyme after dialysis. A. Chromatogram of the products of the enzyme reaction performed with unmodified enzymes. B. Chromatogram of the substrate. C. Chromatogram of the reaction assay performed with di-block polymersomes, incubated for 16h. D. Chromatogram of the reaction assay performed with tri-block polymersomes, incubated for 16h"> <img src="http://openwetware.org/images/5/58/Uplc1t.png" alt="" /> </a> <figcaption>Enzymatic assay</figcaption> </figure> </div>
<div class="clear"></div>
<h2>Smart nanoreactor synthesis</h2>
<div class="project_box" > <p>Until now, we demonstrated that both DNA channel incorporation and enzyme encapsulation were successful. The assembly of DNA channels and encapsulation of active enzyme at the same time are now to be demonstrated.</p>
<h3>Tri-block polymer</h3> <p>In presence of enzyme, the DNA channel incorporation into membrane of polymersome was found to be successful shown using cryo-TEM image. We hypotheses the integration of DNA channels into the polymersome membrane from image contrast modulations around the polymersome membrane which are definitely not present in absence of DNA channel or Enzyme. In addition, the channels were also found to be perpendicular attached to the polymersome surface which may or may not be associated with polymersomes. Further, confirmatory results can be obtained by three dimensional cryo-TEM image collection and reconstruction.</p>
<p>In order to obtain further evidence for DNA channel incorporation into polymersomes, negative stained sample were inspected under TEM. Although one should consider that the polymersomes get at least partly collapsed during this preparation the staining of DNA channels with and without tri-block polymer shows significant morphology differences. In absence of polymer, the DNA channels are clearly visible as cylindrical motif with sharp defined contours. While in presence of polymer, the DNA channels are embedded in a diffuse matrix addressed as polymer shell which is attached to the channel.</p>
<p>In order to confirm our hypothesis, enzymatic analysis was performed and analyzed using UPLC.</p>
<h3>Di-block polymer</h3> <p>As we observed in DNA channel incorporation, the di-block polymersome represented the similar morphology with highly distorted membrane almost present in fragments rather intact polymersomes.</p> </div>
<div class="figure_box"> <figure class="box " > <a class="fb" rel="enzyme_channel" href="http://openwetware.org/images/f/f6/DNAcholp1.png" title="Cryo-TEM images of incorporated DNA channel with cholesterol modification in Tri-block polymersomes in presence of enzymes. The green arrows indicates the dark pattern around polymersome membrane which corresponds to incorporation of the cholesterol-modified DNA channels."> <img src="http://openwetware.org/images/9/93/DNAcholp1t.png" alt="" /> </a> <figcaption>Tri-block DNA-chol</figcaption> </figure>
<figure class="box " > <a class="fb" rel="enzyme_channel" href="http://openwetware.org/images/f/fb/DNApap1.png" title="Cryo-TEM images of incorporated DNA channel with palmitate and archidic modification in Tri-block polymersomes in presence of enzymes. The green arrows indicates the dark pattern around polymersome membrane which corresponds to incorporation of the palmitate and arachidic acid-modified DNA channels."> <img src="http://openwetware.org/images/c/ca/DNApap1t.png" alt="" /> </a> <figcaption>Tri-block DNA-P+A</figcaption> </figure>
<figure class="box " > <a class="fb" rel="enzyme_channel" href="http://openwetware.org/images/9/9a/TemDNA.png" title="TEM images of incorporated DNA channel with cholesterol modifications into the tri-block polymersome."> <img src="http://openwetware.org/images/9/97/TemDNAt.png" alt="" /> </a> <figcaption>TEM Tri-block</figcaption> </figure>
<figure class="box " > <a class="fb" rel="enzyme_channel" href="http://openwetware.org/images/6/6f/DNAcholp2.png" title="Cryo-TEM images of Di-block polymersomes in presence of DNA channel with cholesterol modification and enzyme. The polymersome formation is highly inhibited which can be seen due to aggregation and only few stable polymersome can be seen."> <img src="http://openwetware.org/images/1/1d/DNAcholp2t.png" alt="" /> </a> <figcaption>Di-block DNA-chol</figcaption> </figure>
<figure class="box " > <a class="fb" rel="enzyme_channel" href="http://openwetware.org/images/7/7f/DNApap2.png" title="Cryo-TEM images of Di-block polymersomes in presence of DNA channel with palmitate and archidic modification and enzyme. Morphology of polymersome is highly distorted due to presence of DNA channel and enzyme which can be observed."> <img src="http://openwetware.org/images/6/67/DNApap2t.png" alt="" /> </a> <figcaption>Di-block DNA-P+A</figcaption> </figure> </div> <div class="clear"></div>
<h2>Smart Nanoreactor assay</h2>
<div class="project_box" > <p>The enzymatic assay was performed to prove that our system works: the channel allows the transport of the substrate in and out of the polymersomes, the encapsulated enzyme is still active and the reaction takes place.</p>
<p>The reaction assay was performed after dialysis of the samples, so that no free enzyme is present in the solution. For the samples with DNA channel, two types of reaction were set, one with higher concentration and another with lower concentration of the polymersomes, to distinguish which one is the best to observe the reaction. The reaction was performed and samples were taken at 3h, 8h30min and 48h. Those were frozen and the reaction products were analyzed by UPLC.</p>
<p> For the tri-block polymersomes, conversion of substrate into product could already be observed after 3 h incubation, even in the samples with lower concentration. However, the reaction rate in these samples is slower because even after 48 h the majority of the substrate remained unconverted. Nevertheless, the reaction for the tri-block polymersomes could also be observed in the sample without channels. For this reason, this assay should be repeated with an optimization of the concentration of the enzyme and the dialysis time.</p>
<p>Previous reactions with the polymersomes without channels and the enzyme encapsulated showed that there was no conversion of the substrate, so in this case the apparition of the product could be due to a too short dyalisis time.As a follow-up experiment, proteases will be added outside the polymersomes before the reaction assay in order to make sure that no free enzyme is present in the solution, thus proving that the enzyme inside the nanoreactors is protected and still active.</p>
<p>In the case of di-block polymersomes, the samples showed no reaction, even with higher concentration of polymersomes. This correlates with the previous results that showed that the formation of these polymersomes was affected by the incorporation of the channel, and so no enzyme could be incorporated.</p> </div>
<figure class="box " > <a class="fb" rel="int_rxn" href="http://openwetware.org/images/9/96/Tri-block_negative.png" title="Enzyme reaction assay with tri-block polymersomes without channel. A. Incubation time 3h. B. Incubation time 8h 30 min. C. Incubation time 48 h."> <img src="http://openwetware.org/images/e/e9/Tri-block_negative_thumbs.png" alt="" /> </a> <figcaption>Tri-block without channel</figcaption> </figure>
<figure class="box " > <a class="fb" rel="int_rxn" href="http://openwetware.org/images/c/c5/Triblock_nanoreactors_cholesterol.png" title="Enzyme reaction assay with tri-block polymersomes with channel , having 5’ palmitate and arachidic acid modifications. A. Reaction assay with low concentration of the polymersomes (22 μg/ml).Different incubation times are shown (3h, 8h 30 min, 48h). B. Reaction assay with high concentration of polymersomes (100μg/ml). Different incubation times are shown (3h, 8h 30min, 48h)."> <img src="http://openwetware.org/images/a/a8/Triblock_nanoreactors_cholesterol_thumbs.png" alt="" /> </a> <figcaption>Tri-block channel cholesterol</figcaption> </figure>
<figure class="box " > <a class="fb" rel="int_rxn" href="http://openwetware.org/images/8/84/Triblock_nanoreactors_pa.png" title="Enzyme reaction assay with tri-block polymersomes with channel, having 3’ cholesterol modifications. A. Reaction assay with low concentration of the polymersomes (22 μg/ml).Different incubation times are shown (3h, 8h 30 min, 48h). ). B. Reaction assay with high concentration of polymersomes (100μg/ml). Different incubation times are shown (3h, 8h 30min, 48h)."> <img src="http://openwetware.org/images/7/7b/Triblock_nanoreactors_pa_thumbs.png" alt="" /> </a> <figcaption>Tri-block channel P+A</figcaption> </figure>
<figure class="box " > <a class="fb" rel="int_rxn" href="http://openwetware.org/images/8/8c/Nanoreactors_diblock.PNG" title="Enzyme reaction assay with di-block polymersomes. A. Chromatogram of the substrate. B. Chromatogram of the di-block polymersomes without channel, incubated for 48h with the substrate solution. C. Chromatogram of the di-block polymersomes with channel, having 5’ palmitate and arachidic acid modifications, incubated for 48h with the substrate solution. D. Chromatogram of the di-block polymersomes with channel, having 3’ cholesterol modifications, incubated for 48h with the substrate solution.."> <img src="http://openwetware.org/images/4/48/Nanoreactors_diblock.png" alt="" /> </a> <figcaption>Di-block assay</figcaption> </figure>
<div class="clear"></div>
</section>
<a class="anchor" id="achievements"></a>
<div class="interlude clearfix"><h2>Achievements</h2></div>
<section id="achievements" class="project" data-stellar-background-ratio="0.5">
<h2></h2>
<div class="achievement_box" >
<img src="http://openwetware.org/images/1/1b/Nm_achievement.png" alt="N-acylation reaction" title="">
</div>
<div class="clear"></div>
</section>
<a class="anchor" id="future"></a>
<div class="interlude clearfix"><h2>Future work</h2></div>
<section id="future" class="project" data-stellar-background-ratio="0.5">
<h2></h2>
<div class="project_box" > <p>Our project does not finish here. There is a set of planned experiments that due to the time constraint were impossible to perform. The ones that catch our interest are the following.</p>
<h3>Quantifying the incorporation of the channels</h3>
<p> We will attach gold nanoparticles of around 2-5 nm in size that have attached to their surface a ssDNA sequence that can hybridize with the scaffold of the DNA origami channel.</p>
<p> This will help distinguish the DNA origami channel in the TEM and cryo-TEM imaging and so a quantification of the channels that are present in each polymersome will be made.</p>
<h3>Structural studies of the origami channel</h3>
<p> In collaboration with Prof. Dr. Rasmus Schroeder, in Heidelberg University, we will do structural studies of the DNA origami 3D structure in the nanometer scale.</p>
<h3>Optimization of the Smart Nanoreactor</h3>
<p>The formation of the Smart Nanoreactor will be optimized with regard to the concentration of enzyme, DNA channel and coblock polymers.</p>
<p>Furthermore, reaction could be observed in the negative control. Previously to the reaction assay, proteases will be introduced outside the polymersomes that can degrade the enzymes that are not incorporated. Thus, we will prove that the observed products are due only to the encapsulated enzyme.</p>
<p>In the case of the di-block polymersomes, their formation with the presence of the origami channel was affected. Thus, we will further investigate their formation with lower concentrations of the origami channel.</p>
<h3>Antibody immobilization</h3>
<p>For targeted delivery in the in-vivo studies, the coblock polymers will be functionalized with a suitable linker for the antibody attachment. Thus, we will be able to specifically target cells with our Smart Nanoreactor.</p>
<h3>In-vitro and in-vivo studies</h3>
<p>In-vitro studies with cell cultures will be performed to test the delivery of the antibody-modified nanoreactors to the desired cells.</p>
<p>Furthermore, in collaboration with Prof. Dr. Michael Brand in CRTD, we will perform in-vivo studies using Zebrafish, because due to their transparent body, we will be able to easily trace the Smart Nanocontainer and analyze its efficiency in targeting.</p>
</div> <div class="clear"></div>
</section>
<a class="anchor" id="applications"></a>
<div class="interlude clearfix"><h2>Applications</h2></div>
<section id="applications" class="project" data-stellar-background-ratio="0.5">
<h2>Drug delivery system</h2>
<div class="project_box" >
<p>In recent years DNA origami reached incredible popularity in nanotechnology due to its vast design possibilities and its broad potential applications. While having numerous advantages it is still exceptionally expensive to produce in high amounts for mass applications, whereas inorganic polymers, another range of materials available for biomolecular design can compensate for this disadvantage. Nowadays they can be found in almost every aspect of our lives, in medicine and technology through their wide range of different functionalities and their moderate price. In our project we aim to combine those two artificial materials into one device: functional and responsive polymers as carrier material with highly flexible designable DNA origami for more complex access points to gain the best of both of them.</p>
<p>Our main application presented here in our video we describe using our Smart Nanoreactor as a new form of drug delivery system, which takes great advantages of the polymersome's role as carrier for multiple different other functional components. In chemotherapy a big range of treatment methods use drugs which are damaging both cancerous as well as healthy tissue, putting the patient in a high stress during the treatment. But with out Smart Nanoreactor we offer a solution for this problem. As a first step the Nanoreactor is injected into the patient and through antibodies immobilized on the outside of the sphere, the device is targeting specific pathogenic tissue like cancer and is localized only in this area. In the second step an inactive form of a pro-drug gets systematically introduced into the patient, so it is localized in healthy as well as pathogenic tissue. Now the pro-drug can enter the Nanoreactor through the origami channels and the encapsulated enzyme can convert the inactive pro-drug to its active state in which it is damaging both healthy and pathogenic tissue. After exiting the Nanoreactor again the active drug can interact with its surroundings, due to the location of the Nanoreactor the closest cells are the intended cancerous tissue. After the targeted cells are treated the immobilized antibodies loose their binding partner, the reactor dis-attaches and is filtered out of the body through the kidneys.</p>
</div>
<div class="figure_box">
<figure class="box " >
<a class="fb" rel="application" href="http://openwetware.org/images/9/9d/Comic.png" title="As a first step in the application for a drug delivery system the Smart Nanoreactor is injected into the patient and binds to pathogenic tissue. As a second step a pro-drug is introduced and converted by the Nanoreactor to an active form. Now the drug is able to treat pathogenic cells only.">
<img src="http://openwetware.org/images/9/96/Comic_thumb.png" alt="" />
</a>
<figcaption>Comic</figcaption>
</figure>
<figure class="box " > <a class="fb" rel="application" href="http://openwetware.org/images/5/5d/Nanocontainer.png" title="Immobilizing antibodies on the Nanoreactor enable the targeting of pathogenic tissue like tumors and provide our device with the necessary target specificity to act as a drug delivery system."> <img src="http://openwetware.org/images/5/5f/Nanocontainert.png" alt="" /> </a> <figcaption>Drug delivery system</figcaption> </figure>
</div> <div class="clear"></div>
<h2>pH-responsive Nanocontainer</h2>
<div class="project_box" >
<p>Another powerful feature of our device is the use of a pH-responsive polymer, which enables us to add even more functionality to our Nanoreactor. The described Di-block copolymer has tight and stable walls in basic pH but gets leaky in acidic milieu. We could use this to create another route of transport for molecules that are too big to fit through the origami channel. In high pH the container circles through its environment and takes up small molecules from the outside, an enclosed enzyme crosslinks them together until they are too big to exit again through the channel. After the container reaches surroundings with lower pH, its walls become more porous and allow the exit of its cargo through pores in the polymersome. A great advantage is the reversibility of this process, so when the vesicle is empty, it can be brought back to basic pH, the walls are sealed tight again and the reactor can take up more substrate through the DNA channel.</p>
<p>This feature could not only open a variety of applications in medicine where our device can react to physiological changes in pH but also in technology and environmental engineering for example. Synthesizing processes can be automated and our Nanoreactor can respond to different stages of a pH changing reaction or it could be used to clean freshwater supplies via independent sampling of contaminated wells.</p>
</div>
<div class="figure_box">
<figure class="box " >
<a class="fb" rel="application" href="http://openwetware.org/images/7/70/Nm_pH_response.png" title="The porosity of the pH responsive polymers changes according to pH. This adds even more functionality to our Nanoreactor.">
<img src="http://openwetware.org/images/0/04/Nm_thumb_pH.png" alt="" />
</a>
<figcaption>pH responsive Nanocontainer</figcaption>
</figure>
</div>
<div class="clear"></div>
<h2>Smart Nanofactory</h2>
<div class="project_box" >
<p>The possibilities of our device for technological applications do not end here but observing nature and discovering the formation of polymersomes inside bigger polymersomes during Cryo-TEM analysis another great range of technical applications came to our minds. If we extend this process to creating complete Smart Nanoreactors encapsulated in other Nanoreactors we can create whole Nanofactories in the future, similar to the different lipid vesicle compartments in living cells like vacuoles.</p>
<p>Combining different reactors with various different DNA origami channels, polymersomes and enzymes we are able to design a system, which can independently catalyze complex chains of reactions. Immobilized targeting proteins on the outside of polymersomes and their sizes act for sorting of the reactors, while the different channels determine the order of entry that the substrates have to use to complete the full chain of a reaction. Besides easy separation of product molecules form enzymes, our device has the advantage of formation of big structures by different enzymes in a complex order, one can even create repeats and loops by incorporating a range or Nanoreactors within each other, all of it independently.</p>
<p>This was just a small trip through the wide spectrum of applications for our Smart Nanoreactor, but there is far more possible in the future. And all of this just with a plastic ball.</p>
</div>
<div class="figure_box"> <figure class="box " > <a class="fb" rel="application" href="http://openwetware.org/images/a/ac/Nanofactory_1.png" title="Combining different reactors with various different DNA origami channels, polymersomes and enzymes we are able to design a system, which can independently catalyze complex chains of reactions."> <img src="http://openwetware.org/images/4/44/Nanofactory_thumb.png" alt="" /> </a> <figcaption>Nanofactory</figcaption> </figure>
</div>
<div class="clear"></div>
</section>
<a class="anchor" id="references"></a>
<div class="interlude clearfix"><h2>References </h2></div>
<section id="references" class="project" data-stellar-background-ratio="0.5">
<div class="clear"></div>
<h2></h2>
<div class="project_box refernces">
<ul> <li> Ben-Haim, N. et al., Cell-Specific Integration of Artificial Organelles Based on Functionalized Polymer Vesicles, Nano Lett. 8 (2008) 1368-1373 </li> <li> Blanazs, A. et al., Self-Assembled Block Copolymer Aggregates: From Micelles to Vesicles and their Biological Applications, Macromol. Rapid Commun. 30 (2009) 267–277 </li> <li> Bonroy, K. et al., Comparison of random and oriented immobilisation of antibody fragments on mixed self-assembled monolayers, Journal of Immunological Methods 312 (2006) 167–181 </li> <li> Broz, P. et al., Cell targeting by a generic receptor-targeted polymer nanocontainer platform, Journal of Controlled Release 102 (2005) 475–488 </li> <li> Broz, P. et al., Toward Intelligent Nanosize Bioreactors: A pH-Switchable, Channel-Equipped, Functional Polymer Nanocontainer, Nano Lett. 6 (2006) 2349-2353 </li> <li> Canthon, Ir. et al., Fully synthetic polymer vesicles for intracellular delivery of antibodies in live cells, FASEB Journal 27 (2013) 1-11 </li> <li> Checot, F. et al., pH-responsive micelles and vesicles nanocapsules based on polypeptide diblock copolymers, Biomolecular Engineering 24 (2007) 81–85 </li> <li> Christian, D.A. et al., Spotted vesicles, striped micelles and Janus assemblies induced by ligand binding, Nature Materials 8 (2009) 843-849 </li> <li> van Dongen, S.F.M. et al., Cellular Integration of an Enzyme-Loaded Polymersome Nanoreactor, Angew. Chem. Int. Ed. 49 (2010) 7213 –7216 </li> <li> Douglas, S.M. et al., Rapid prototyping of 3D DNA-origami shapes with caDNAno, Nucleic Acids Research 2009 1–6 </li> <li> Egli, S. et al., Biocompatible Functionalization of Polymersome Surfaces: A New Approach to Surface Immobilization and Cell Targeting, J. Am. Chem. Soc. 133 (2011) 4476–4483 </li> <li> Gaiduk, A. et al., Making gold nanoparticles fluorescent for simultaneous absorption and fluorescence detection on the single particle level, Phys. Chem. Chem. Phys. 13 (2011) 149–153 </li> <li> Le, H.T. et al., Antibody functionalization with a dual reactive hydrazide/click crosslinker, Analytical Biochemistry 435 (2013) 68–73 </li> <li> LoPresti, C. et al., Polymersomes: nature inspired nanometer sized compartments, J. Mater. Chem. 19 (2009) 3576–3590 </li> <li> Luxenhofer, R. et al., Doubly amphiphilic poly(2-oxazoline)s as high-capacity delivery systems for hydrophobic drugs, Biomaterials 31 (2010) 4972-4979 </li> <li> Mabrouk, El. et al., Bursting of sensitive polymersomes induced by curling, J. Am. Chem. Soc. 18 (2009) 106 </li> <li> Maglia, G. et al., DNA Strands from Denatured Duplexes are Translocated through Engineered Protein Nanopores at Alkaline pH, Nano Lett. 9 (2009) 3831-3836 </li> <li> Marguet, M. et al., Multicompartmentalized polymeric systems: towards biomimetic cellular structure and function, Chem. Soc. Rev. 42 (2013) 512-529 </li> <li> Meeuwissen, S.A. et al., Copper-free click chemistry on polymersomes: pre- vs. post-self-assembly functionalisation, Polym. Chem. 3 (2012) 1783–1795 </li> <li> Meng, F. et al., Polymersomes Spanning from Nano- to Microscales: Advanced Vehicles for Controlled Drug Delivery and Robust Vesicles for Virus and Cell Mimicking, J. Phys. Chem. Lett. 2 (2011) 1533–1539 </li> <li> Nardin, C. et al., Giant Free-Standing ABA Triblock Copolymer Membranes, Langmuir 16 (2000) 7708-7712 </li> <li> Tanner, P. et al., Enzymatic Cascade Reactions inside Polymeric Nanocontainers: A Means to Combat Oxidative Stress, Chem. Eur. J. 2011 (17) 4552–4560 </li> <li> Onaca, Oz. et al., Functionalized Nanocompartments (Synthosomes) with a Reduction-Triggered Release System, Angew. Chem. Int. Ed. 47 (2008) 7029 –7031 </li> <li> Opsteen, J.A. et al., ‘‘Clickable’’ polymersomes, Chem. Commun. (2007) 3136–3138 </li> <li> Renggli, K. et al., Selective and Responsive Nanoreactors, Adv. Funct. Mater. 21 (2011) 1241–1259 </li> <li> Rooseboom, M. et al., Enzyme-Catalyzed Activation of Anticancer Prodrugs, Pharmalogical reviews 56 (2004) 53-102 </li> <li> Rothemund, P.W.K., Folding DNA to create nanoscale shapes and patterns, Nature 440 (2006) 297-302 </li> <li> Sauer, M. et al., Polymer Nanocontainers for Drug Delivery, American Chemical Society 16 (2004) 224-237 </li> <li> Sjoeback, R. et al., Absorption and fluorescence properties of fluorescein, Spectrochimica Acta Part A 51 (1995) L7-L21 </li> <li> Stoenescu, R. et al., Asymmetric ABC-Triblock Copolymer Membranes Induce a Directed Insertion of Membrane Proteinsa, Macromol. Biosci. 4 (2004) 930–935 </li> <li> Wei, X. et al., Mapping the Thermal Behavior of DNA Origami Nanostructures, J. Am. Chem. Soc. 135 (2013) 6165-6176 </li> <li> Xu, G. et al., Strategies for Enzyme/Prodrug Cancer Therapy, Clin Cancer Res 7 (2001) 3314–3324 </li> <li> Yassin, M.A. et al., pH-Dependent Release of Doxorubicin from Fast Photo-Cross-Linkable Polymersomes Based on Benzophenone Units, Chem. Eur. J. 18 (2012) 12227 – 12231 </li> <li> Zhang, X.-B. et al., Metal Ion Sensors Based on DNAzymes and Related DNA Molecules, Annual Review of Analytical Chemistry 4 (2011) 105-128 </li> </ul>
</div>
<div class="clear"></div>
</section>
</div>
</section> <!-- #content --> <script>
$(function () { $('body').stellar(); $(".fb").fancybox({ openEffect : 'elastic', closeEffect : 'elastic', helpers : { title : { type : 'inside' }, overlay: { locked: false } } }); $.scrollUp({ scrollText: '', // Text for element, can contain HTML scrollTitle: true, // Set a custom <a> title if required. Defaults to scrollText scrollImg: true, // Set true to use image zIndex: 2147483647 // Z-Index for the overlay }); $("#subnav").sticky({ topSpacing: 200, bottomSpacing:100 }); $.localScroll(); }); </script> </body> </html>